Human brain theory
ISBN
978-3-00-068559-0
Monograph of Dr. rer. nat. Andreas Heinrich Malczan
11. The further development of the ladder nervous system in humans
In this monograph, it has often been shown that the brain's signal processing is realised by modules. These often produce as output submodalities that have arisen through the splitting of original modalities. For example, the new submodality of grey values emerged from the modalities light and dark. From the modalities green and red emerged the submodality of the mixed colours red/green, likewise a submodality emerged for the mixed colours red/blue and blue/green. We can combine these three submodalities into the new submodality of colour, since the three modules in the brain are exactly superimposed per image pixel, including the brightness module.
In the divergence module with spatial signal propagation, the new modality of oriented line elements emerged, both for the submodality of colour and for the submodality of grey values. From it, in interaction with the basal ganglia, emerged the modality of the moving line elements for both colours and greys.
In the human brain, but probably much earlier in the vertebrate nervous system, a tendency developed to separate different modalities. In the course of evolution, this was also transferred to the submodalities. They represented new, previously unknown signals.
Of course, these new signals were not generated by receptors, but they were nevertheless the output of neurons in the various modules of the rope ladder nervous system or the successor system.
We remember: Most modules - for example in the visual field - were created in the process of signal divergence. Output was distributed to several, later to many neurons. This served to increase fail-safety, represented a kind of formation of replacement neurons. It is nonsensical to assume that this only happened in the cortex of higher vertebrates. No, this process may have begun very early in evolution. Slowly, almost insidiously, the number of output neurons in the sensory and motor centres was simply increased. Together with the increase in the types of receptors and the number of receptors, this led to a spatial expansion in the head segments. The head skeleton there - once present - probably acquired the ability to simply expand as the number of neurons in its interior increased. Even today, the fontanelles in newborns allow the brain to expand during the first two years of life. We assume here that this cephalic skeleton was already present.
As the layer thickness of the sensory and motor centres slowly grew, which also automatically created the divergence modules as a result of signal divergence, there was new output from the resulting submodalities (brightness/colour/orientation of lines, etc.).
In those segmented bilateria whose lineages led to primates and Homo sapiens, a completely new, unprecedented concept began to take hold:
The output of the new submodalities from the different modules of the first floor was equated to the receptor output.
It was treated as if a new type of receptor had emerged that delivered this output in the uppermost conductor segment.
This had far-reaching consequences. In the ladder system of the head segments, each modality claimed its own ladder segment. When the new submodalities developed, they also had to claim their own conductor segment. Thus, a new conductor segment for the signals of the new submodalities was created above the previous uppermost, cortical conductor segment. A cortex above the cortex, so to speak. The signals of the new submodalities were passed on upwards.
Technically, however, this was difficult to implement. No new conductor segment could be created at that height; the available space was completely filled by the existing neurons and axons and was already limited at the top by the skull bone (even though it may still have consisted of cartilage at that time).
Thus, the only option left for the new ladder segments was to move to the side. The next ladder segment therefore formed laterally next to the cortical ladder segment. Between the previous cortical segment and the new segment, however, there were the classical connections.
Perhaps this development was caused by a peculiarity of neuron class 4. In the course of evolution, they specialised in transmitting new signals to the next neighbouring segment. If there was no superior segment, they created a new one themselves by having their axons follow the existing gradient gradient of an existing head marker and activating existing class 3 proneurons so that these became class 3 projection neurons. Their axons - controlled by the motor marker of the body side in question - moved to the motor side and generated new class 5 projection neurons, which in turn projected downwards. This created a new floor, i.e. a new segment in the nervous system.
It was the same process as the emergence of new receptors. Their signals ultimately also generated the class 4 neurons in the respective segment, which passed them on to the higher-level segments. The only new thing was that in the case where there was no higher segment, one was newly created.
The sensory centre of the previous cortical segment was connected to the sensory centre of the new segment via class 4 neurons. Their axons transported the signals of the new submodality from the previous cortical divergence module to the higher-level sensory centre of the new segment. Similarly, the motor centre of the cortical segment was connected to the motor centre of the new superior segment.
And finally, in this new segment, the sensory centre was also linked to the motor centre via class 3 neurons. At the same time, the new segment projected via class 2 neurons into lateral exchange nuclei, which projected to the opposite side and made contralateral inhibition possible there.
The mean centres also received (from a certain evolutionary stage) the signals of this new conductor segment and caused a back projection to it.
Thus, a secondary cortex developed alongside the primary cortex, which received input from it and projected back to it. We can also call this secondary cortex the association cortex because - for the first time in evolution - it no longer received direct receptor signals, but the signals of the submodalities from the divergence modules (and other modulars) of the primary cortex. And these were treated as if they were receptor signals.
We illustrate the gradual emergence of this secondary cortex in a figure relating to the ladder system. It shows how new ladder segments always emerged above the primary, cortical ladder segment, and how they each attached themselves laterally, whereby all axons now had to run on semi-circular paths within the white matter.
Theoretically, these conductor
modules could also be arranged classically in the form of a single conductor,
were it not for the skullcap that prevents this.
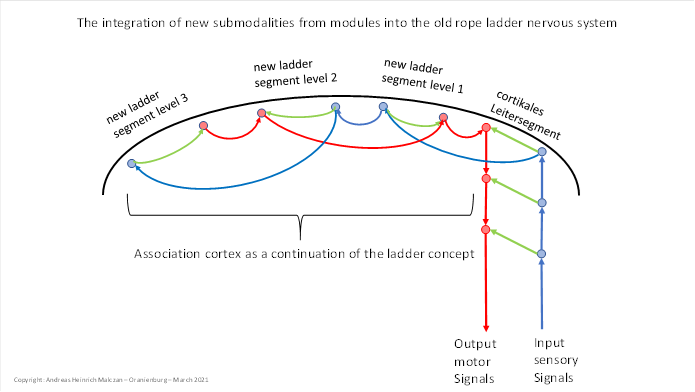
Figure 51: The integration of new submodalities into the old rope ladder nervous system
One consequence of this lateral accumulation of the newly formed conductor segments was the lateral expansion of the structures. Today, we commonly refer to them in humans, but also in vertebrates, as cortex. We can retain this linguistic regulation, but we should always remember that the original cortex represented the uppermost conductor segment, which still received its input via classical receptors.
Now, even after the topmost, cortical ladder split into four different subladder of different modality groups - the lobi of the brain emerged from this - each of these subladder began a lateral growth to accommodate the newly emerging, higher-level ladder segments for the self-generated submodalities.
We thus record lateral cortical growth not only through signal divergence in the area, but through the emergence of submodalities that gave rise to an association cortex.
The emergence of this association cortex laterally from the existing, primary cortex took place in all lobes of the brain and on both sides of the body. In each lobe of the brain, the (quite early onset of) signal divergence gave rise to the divergence modules, whose mode of operation was the same everywhere, but whose input differed. This is because quite early on, the receptors had already come together in subclasses, each of which claimed a future brain lobe for itself.
The extent to which this association cortex or its precursor, the association ladder, already developed in the vertebrates of the early evolutionary period will not be discussed further here. Certainly, the association cortex of modern humans developed gradually from the early rope ladder nervous system.
This also explains the connections between the cortex fields. One always finds sensory and motor sub-areas alternating in the association cortex, which are linked to each other via neurons of class 4, class 3 and class 5. Also, the class 6 and class 1 mean neurons - omitted in the figure above - continue to exist in the association areas as well. Likewise, the areas of the association cortex project into the opposite side via class 2 neurons, whose projection axons form the structure of the bar in the brain. Via projection into the dopaminergic substantia nigra pars compacta and back projection to the striatum, these new brain structures are also integrated into the basal ganglia system. Furthermore, the cerebellum receives the new elementary signals via the bridge nuclei, while the mean projection ends at the climbing fibres of the pontocerebellum. Thus, new complex signals can again be derived from the signals of these new association areas. Feeding into the limbic system also takes place as before.
We owe our intelligence (among other things) to three principles that became established in the course of the evolution of the central nervous systems:
As far as possible, every signal was taken into account in the mean kernels. This also applied to signals that had only arisen through the work of neuronal modules and thus represented new submodalities.
Every new signal that arose in one floor of the segmented nervous system through the work of neuronal modules was passed on to the higher floors.
If there was no higher floor for it, the projection neurons of class 4 established a new floor, so that a superior neuronal segment was created. This had to move sideways due to the limitation by the skullcap. This is how the secondary association cortex was formed.
Possibly, the head skeleton could also grow in height in some species, so that new floors of the association system were simply added on top there. This lengthened the head in the shape of a piston.
Monograph of Dr. rer. nat. Andreas Heinrich Malczan