Human brain theory
for the European Union’s Human Brain Project
ISBN 978-3-00-068559-0
9. The inverse pontocerebellum
As early as 2012, I developed the concept of the inverse cerebellum. One reason for this was the principle I discovered that any new signal form produced by any module must always be transformed back into the original signal form so that it can perform its previous tasks in the system.
If the pontocerebellum now transforms the elementary signals into complex signals, another module must transform these complex signals back into the original elementary signals. Otherwise, these would simply be lost forever in the pontocerebellum instead of causing their previous effects.
Since the complex signal to an active neuron population of elementary signals is formed by the pontocerebellum, the theoretically necessary inverse structure was initially given the name "inverse cerebellum".
The mode of operation was clear in principle. The pontocerebellum formed the new class of complex signals from elementary signals. For this, it needed the mean climbing fibre signals for the imprinting of the Purkinje cells by means of LTD and LTP. The elementary signals were the mossy fibre input. The output of the pontocerebellum was the complex signals.
These had to be transformed back into elementary signals in the inverse pontocerebellum.
The original elementary signals travelled from the (primary) cortex (among others) to the nucleus ruber. In this way they (also) reached the side-changing nucleus of this floor, the nucleus olivaris. From there they travelled as climbing fibres to the pontocerebellum.
Now, a timed climbing fibre signal is required for the imprinting process, so it must be interrupted by short pauses. In these pauses, Purkinje groups recognise their own signal and inhibit the climbing fibre signals of the olive so that no multiple imprinting occurs.
The short pauses were accomplished by the basal ganglia system. It also received the same elementary signals, caused a short time delay and projected inhibitory signals into the (ventral) thalamus, where they interrupted the original elementary signals. Since this also interrupted the signal flow to the cortex and thus also to the basal ganglia, the inhibition of the elementary signals fell away with a certain time delay. However, all this only applied to stationary signals, i.e. those that had a sufficiently long signal duration. For short-lived signals, the basal ganglia effected movement detection by inhibiting the time-delayed echo so that no residual signal remained from unchanged signals. In this way, movements and signal changes became recognisable.
With stationary (unchanging signals with a longer signal duration) it was different. Here, the short pauses that were absolutely necessary for imprinting in the pontocerebellum were imposed on the elementary signals.
Each elementary signal thus activated not only the mossy fibres in the pontocerebellum, but also (at least) one climbing fibre in the inverse pontocerebellum. We recall that (according to this theory) every cortical elementary signal already moved in early times via the nucleus ruber and the nucleus olivaris to the opposite side and reached the cerebellum there as a climbing fibre. Now it only needed to move on and generate its own Purkinje cells in the midline area, the pontocerebellum.
By imprinting, these Purkinje cells now learned all complex signals that were active during the activity of this elementary signal. Due to a signal divergence of the climbing fibres (in the nucleus olivaris), Purkinje cells in different segments of the inverse pontocerebellum were also able to include complex signals in their imprinting that had exactly this elementary signal in their signal spectrum. This is because many elementary signals were present as signal components in different complex signals.
Whenever a complex signal was active whose imprinting set included a certain elementary signal, exactly the Purkinje cell in the inverse cerebellum that was assigned to this elementary signal was activated.
Thus, the inverse pontocerebellum transferred all complex signals back into the associated elementary signals, whereby the imprinting, i.e. a learning process, was necessary for this. Only after this process was completed could a complex signal be transferred back into the elementary signals.
The output of the inverse pontocerebellum took the path to the thalamus and contacted exactly those neurons there that were assigned to the elementary signals.
If the activity of a certain neuron population in the thalamus led to the excitation of the associated elementary neurons in the cortex, these signals reached the associated Purkinje cell via the bridge nuclei and the moss fibres. The latter recognised this signal as its own signal and activated a complex neuron in the secondary cortex.
This complex neuron activates an assigned population of Purkinje groups in the inverse pontocerebellum, each of which is assigned to one of the active elementary signals. The output of the inverse pontocerebellum now activates in the thalamus exactly the population of neurons associated with this complex signal.
This creates a feedback loop that constantly reactivates the active complex signal and its associated elementary signals. The signal oscillates with a constant change of its signal form from the elementary form to the complex form and back again to the elementary form.
This signal oscillation has been demonstrated in the human brain in real terms (e.g. by Wolf Singer).
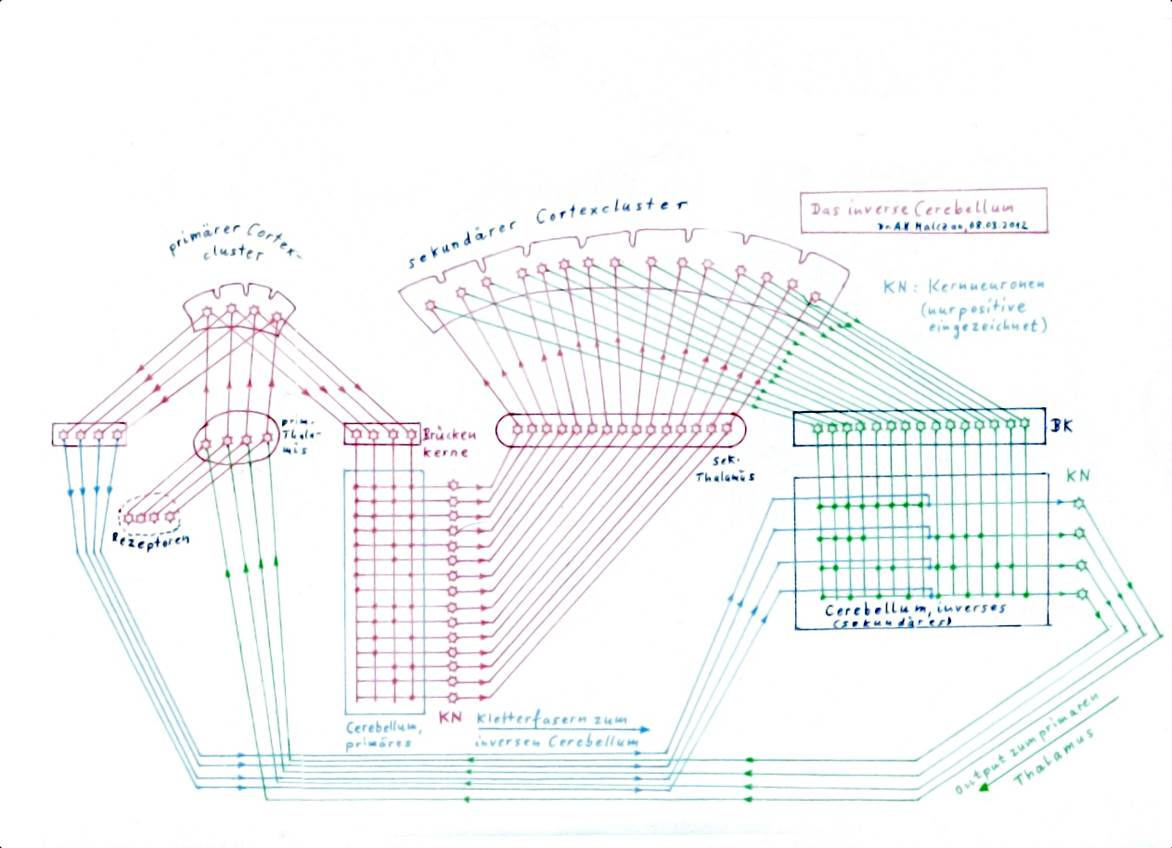
Figure 48: Cerebellum and inverse cerebellum - basic circuitry
In the figure above, the path of the cortical elementary signals to the inverse cerebellum is highly simplified. In reality, it consists of several stages and two parallel pathways. The cortex projects via the nucleus ruber to the nucleus olivaris, which in turn sends the climbing fibres to the inverse cerebellum (shown in green above). On the other hand, the cortex projects to the substantia nigra, which projects back to the striatum, which in turn projects inhibitively to the thalamus, where it causes a brief signal interruption in the case of stationary signals. This signal interruption is passed on via the cortex to the climbing fibre projection to the inverse cerebellum. In this way, the climbing fibre signals also acquire their typical form in the inverse cerebellum, an oscillation that is interrupted by short pauses. It is ideally suited for the imprinting of Purkinje cells.
What would be the point of a complex signal activating all its elementary signal components? After all, the elementary signals assigned to it are active anyway when the complex signal is active.
The reason for this is that incomplete signals are also recognised in the pontocerebellum. If, for example, 200 elementary signals belong to a certain complex signal, the associated dentate neuron will also provide an output signal if, for example, only 150, 120 or even only 100 of these 200 elementary signals are active.
The dentate neuron then reports the recognition of this complex signal to the secondary cortex, although only just under half of the necessary elementary signals were active. If the inverse pontocerebellum receives this complex signal as input and completely activates all the associated elementary signals, then all 200 elementary signals are suddenly activated in addition to the 150, 120 or even only 100 already active elementary signals. As a result, 50, 80 or even 100 elementary signals are now suddenly additionally active, which were previously completely inactive.
Thus, the inverse cerebellum supplements incomplete signals at the level of elementary signals. This signal supplementation represents the brain's self-created input, which - at higher levels - we may well regard as thoughts, intuitions, ideas, etc. They arise from the subconscious, whose material seat must be located in the cerebellum, and reach consciousness in the cortex. They arise from the subconscious, whose material seat must be located in the cerebellum, and reach consciousness in the cortex.
If the question "Who eats oats" is asked, the inverse pontocerebellum completes the answer because the complex signal "Horses eat oats" is activated, even if it is only incomplete. It is activated because it has the partial signals "oats" and "eat". Only the partial signal "oats" is missing. Nevertheless, it is sufficient for the activation of this complex signal. The complex signal "horses eat oats" in turn activates the signal components (elementary signals) "horses", "oats" and "eat" in the inverse pontocerebellum. Since the signals "oats" and "eat" were already components of the question, they are active and suppress the output of the inverse pontocerebellum. This leaves only the signal "horses" as the answer. Thus the answer is found: "Who eats oats? Horses!
Without the inverse pontocerebellum, the complex signals of the pontocerebellum would remain stuck in it, there would be no algorithm that produces self-created signals. In this respect, we owe our thinking predominantly - but not only - to the interaction of the pontocerebellum and the inverse pontocerebellum.
All this is made possible by the feedback between the pontocerebellum and the inverse pontocerebellum. And since both can certainly be interpreted as (modified) neural networks, the supporters of feedback neural networks will feel confirmed in their assumption that only meaningful feedback can produce artificial intelligence that is capable of more than just pattern recognition.
I already formulated this insight in my first monograph:
Theorem about the emergence of consciousness in the brain
As the number of signals in the recursively structured system from the cerebellum clusters of the direct and inverse pontocerebellum increases, the inhibitory strength in the receptive neighbour inhibition of the thalamus decreases with increasing number of steps of the recursions due to distance.
In this way, the output of the inverse systems can become independent and, in superimposition and feedback with the current as well as with the earlier, cached input, create an inner, multimodal and time-varying image of the world, which can be called consciousness.
The difference between classical neural networks and the cerebellum is the dichotomy of the input: The elementary signals come via the moss fibre system, they are the actual input of the pontocerebellum. The mean signals for the pontocerebellum or the elementary signals for the inverse pontocerebellum are the write signals, i.e. the write commands that lead to the storage of the current signal combination of the data lines in the Purkinje cells. These write commands do not exist in classical neural networks, although Google is now also accelerating the imprinting of artificial neurons in the neural network with certain algorithms. And a low-frequency write signal that then acts as an erase signal does not exist in the classical neural networks.
A target function and a comparison with it also do not exist in the cerebellum. Nevertheless, the system is self-learning. If it is fed with "bullshit" input, it will also produce bullshit. Examples are the followers of conspiracy theories who live in their own information bubbles and reinforce their own views.
A large part of the learned knowledge is also no longer learned through one's own "experience", but is passed on via instruction and acquired from many sources. In this respect, artificial intelligence would also be exposed to these influences and could easily be misused. Incidentally, the emergence of an individual will in such artificial systems, which would then no longer be externally controllable, is completely unexplored.
There are other system components in the human brain besides the cerebellum and their inverse subsystems that contribute to intelligence, but they are not presented in this monograph. The reason is the lack of response to my previous efforts to put the workings of the brain on solid scientific footing. It seems the great interest in the workings of the human brain is more often just faked to get generous funding while keeping the scientific competition down. Moreover, there also seem to be dogmas for whose defence one is even willing to sacrifice the freedom of research. Those who swim against the tide may be right, but it will do them little good. Often, adherents of a common view have to die out before new avenues of thought are allowed. One such view is that one cannot know how the brain works. Although unproven, people seem to believe that the brain cannot recognise itself. Here one forgets that the self-recognising brain is not capable of its own recognition out of itself, but that it merely applies the knowledge acquired over thousands of years by hundreds of thousands of scientists in order to recognise connections that have hitherto been neglected. Paying attention to the non-linear signal attenuation of neuronal signals and the resulting consequences for the superposition of such signals would give those responsible for the Human Brain Project a means of understanding the principle of extreme value coding in the human and vertebrate brain and thus open up a new approach to understanding how it works. May my words not go unheard!
Monograph of Dr. rer. nat. Andreas Heinrich Malczan