Signal Theory of Intelligence
for the European Union’s Human Brain Project
15 The transition to digital signal processing in the brain
It is often emphasised that the brain differs fundamentally from a computer. However, this comparison is misleading. At first glance, the crucial difference appears to lie in the digital mode of operation of computers. Yet the brain also processes digital signals – and to a far greater extent than is commonly assumed. Only a small proportion of neural activity is actually analogue. To understand the further development of biological neural networks, it is therefore necessary to examine the mechanisms of digitalisation in the brain more closely. The digitalisation of neural signals is based on the differentiation of receptors into on-off types, the non-linear signal attenuation in axons, and the superposition of neural signals in divergence modules.
15.1The differentiation of receptors into on-off types
The path to the digitisation of analogue firing rates began with the functional differentiation of receptors. From receptors that were initially exclusively ‘on-like’ – whose firing rate increased with increasing signal strength – receptors developed whose firing rate decreased with the strength of the measured physical quantity. On-receptors exhibit a strictly , monotonically increasing response function, whilst off-receptors exhibit a strictly monotonically decreasing one. In the visual system, both types exist for both brightness and colour perception. An ‘on’ signal can be converted into an ‘off’ signal through inversion by inhibiting a sustained signal. In visual receptors, this signal inversion arises through the inhibition of a continuous output signal generated by so-called band synapses.
Where evolution did not produce off-receptors, the off-signals were formed by signal inversion in the spinocerebellum.
The basis of the secondary brain system is the separation of on and off signals. Receptors convert the strength of a primary quantity into a firing rate:
- On signal: firing rate increases with increasing signal strength.
- Off signal: firing rate decreases as signal strength increases.
From what was originally a single type of receptor, differentiation gave rise to two types of receptor, one of the ‘on’ type and the other of the ‘off’ type. This also led to the separation of the signalling pathways, resulting in the formation of double layers.
15.2 On-off signals and Malczan’s double-layer theory
The basis of the secondary brain system is the separation of on and off signals. Receptors convert the strength of a primary quantity into a firing rate:
- On signal: firing rate increases with increasing signal strength.
- Off signal: Firing rate decreases as signal strength increases.
This duality leads to the formation of double layers in the nervous system:
- Each double layer represents a modality (e.g. flexor muscles vs. extensor muscles).
- In the olivary nucleus, such double layers form for joint signals
- Cortical double layers also developed via the cortical projection.
- In the visual system, these double layers are found, for example, in the visual thalamus and (somewhat hidden) in the primary visual cortex, where the input layers are arranged into on-off sublayers. Such a separation can also be assumed for the projection to the cerebellum.
- With the separation of receptors into on and off signals, a separation of marker substances into on and off types also arose, which were necessary for the organisation of the topology of the double layers.
- Via the olivary nucleus, these two marker types were transmitted to the cerebellum, where they led to the striped patterning of the cerebellar cortex.
Evolutionary significance:
- The double-layer organisation arose from the co-activation of motor antagonists.
- Experimental findings (e.g. Zhou et al. 2024) confirm the existence of such marker structures in the cerebellum.
Reference
Andreas Malczan: The double-layer theory according to Andreas Malczan, Chapter 17 in Human Brain Theory (2024/2025), gehirntheorie.de.
15.3 Signal divergence to increase reliability
Nerve cells are living structures. Their functioning can be impaired or they may even die as a result of a wide variety of influences. It was therefore advantageous for the nervous system to transmit important signals via several pathways simultaneously and in parallel. This increased reliability and ensured the stability of information processing. Various forms of signal divergence developed.
Forms of signal divergence:
- Linear divergence: An axon branches along its length. Other neurons tap into these branches, so that the excitation is distributed across several output neurons.
- Spatial divergence: Axon bundles or interneurons distribute the excitation across a plane. Many output neurons access these distributed signals, causing the input to be spread across an area.
- Spatial divergence: The excitation spreads out in three dimensions and is picked up by different neurons in different regions.
One consequence of signal divergence was signal attenuation depending on the path length of the signals.
15.4 Signal attenuation and path length
Signals are transmitted along axons. Constantly open ion channels along the axons lead to changes in ion concentration within the axon as ions flow in or out. Consequently, the excitation decreases with increasing distance, so that only a fraction of the original neuronal activity reaches the output neuron.
The decrease in excitation as a function of distance is referred to here as the transfer function:
- Along a single axon, it is exponential and strictly concave, in accordance with the classical cable equation (developed and measured, amongst others, by Wilfrid Rall in the 1960s):
![]()
- In the plane, the attenuation is greater, as the area increases quadratically with the diameter; the transfer function here is strictly convex:
![]()
- In three-dimensional propagation, the transfer function is in principle identical to that in the plane, as one can imagine bending all the axons so that they lie in a single plane again.
15.5 Signal superposition in divergence modules
In divergence modules, neural excitations from multiple signals superimpose additively.
- When multiple excitations from different locations converge, their transfer functions superimpose.
- These superpositions always have a global extreme value:
- Linear divergence modules → signal minimum.
- Flat divergence modules → signal maximum.
- Spatial divergence modules → signal maximum
- The signal minimum of the linear modules is inverted in the spinocerebellum and converted into a maximum-coded signal. The spinocerebellum acts here as an inversion module.
- The global extreme value uniquely encodes the value of the receptor’s original quantity via the signal strength ratio of the on-off signal pair.
Example of a linear divergence module
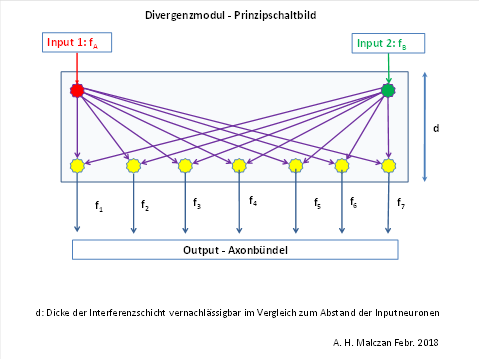
In a planar divergence module, the input neurons are arranged, for example, at the corners of a square, whilst the output neurons are distributed approximately uniformly within this area. Each output neuron is synaptically connected to every input neuron.
15.6 Lateral inhibition and Gaussian encoding
The emergence of a maximum-encoded transfer function in divergence modules or in the subsequent inversion module enables the brain to transform analogue signals into Gaussian encoding. In this process, a continuous firing rate is mapped onto a discrete scale, whereby the neuron with the maximum excitation partially suppresses the excitation of the remaining neurons through lateral inhibition. This leaves a bell-shaped excitation curve, the maximum of which represents the signal strength.
- The neuron whose membrane potential reaches the maximum through temporal and spatial summation of the inputs is the first to reach the threshold for an action potential.
- This neuron fires first and immediately activates the inhibitory interneurons.
- The inhibitory interneurons suppress the only partially amplified excitation of all competing neurons.
Thus, the well-known principle of neuronal competition applies:
“The winner takes all.”
Consequences for signal processing:
- The maximum-coded signal vector in the efferent signal pathway becomes a sparsely coded vector.
- Only a few vector positions have a value greater than zero. At all other positions, the excitation is limited by a kind of Gaussian curve. Advantage: Continuity of the mapping
- This corresponds to classical Gaussian coding.
- Should the currently most excited neuron fail, a neighbouring neuron takes over the signal maximum.
- This makes neural circuits extremely fail-safe and, at the same time, energy-efficient.
The conversion of analogue firing rates into a vector with Gaussian coding represents the digitisation method of the vertebrate nervous system. These vectors of a receptor’s analogue firing rate are almost orthogonal to one another and ideally suited for signal analysis in a wide variety of neural networks such as KNNs, CNNs or RNNs. They also form the signal basis for the emergence of the highest class of networks, the transformers.
15.7 The digital evolution of the basal ganglia
Early vertebrate brains operated exclusively with analogue signals. The differentiation required for motion detection arose in the striatum, which originally consisted solely of striosomes. These contained GABAergic projection neurons that received motor output signals from the dopaminergic medium nucleus. Two signal pathways of different lengths – one excitatory, one inhibitory – enabled the cancellation of time-invariant signals and the amplification of temporal changes. This was the nervous system’s first analogue differential circuit.
As motor function became increasingly complex, analogue differentiation was no longer sufficient. Motor decisions had to be made not only gradually but also categorically. This led to the digitisation of the signals: analogue input values were converted into discrete, one-hot-like signal vectors. To process these new digital representations, the striatum was expanded. Alongside the striosomes, the matrix zones emerged, which form the basis of today’s Go/No-Go circuitry. Thus, the basal ganglia transformed from an analogue difference detector into a digital decision-making module that selects categorical motor options and optimises them on the basis of reinforcement.
15.8 Exciting Recurrence: Quadruple Signal Inversion as a Prerequisite for RNN Functionality
Recurrent neural networks require excitatory signal echoes to stabilise, integrate and recognise temporal patterns. However, the early basal ganglia generated exclusively inhibitory echoes. In order for these signals to be fed into recurrent cortical networks, the brain had to find a solution to convert the inhibitory, delayed signals back into excitatory, maximum-coded signals.
This was achieved through an evolutionarily developed four-fold signal inversion spanning several core regions:
First inversion – striatum
Inhibition of a continuous signal produces the necessary reversal of monotony, but results in a minimally encoded representation. GABAergic projection neurons with large dendritic fields received a kind of sustained excitation from the passing cortical pyramidal cells. From the substantia nigra pars compacta, these received a copy of the cortical signals, which inhibited the striatal neurons. This was the first inversion of the cortical signals.
Second inversion – Globus pallidus externa
The originally unified pallidal system split functionally. The GPe inverts the minimally encoded signal once again to restore the original form of encoding, though it remains inhibitory and delayed. It is required for motion analysis via a difference circuit. Here, the time-delayed signal inhibits the corresponding current signal in the thalamus. The continuous signal for the inversion is supplied by the subthalamic nucleus.
Third inversion – globus pallidus interna
The GPi performs a further inversion; here too, the subthalamic nucleus supplies the necessary continuous signal for the inversion.
Fourth inversion – Thalamus
The final inversion takes place at an excitatory output neuron in the thalamic median nucleus. It is only here that the signal becomes excitatory again, is encoded at maximum amplitude, and remains time-delayed at the same time.
Following this four-stage transformation, the signals can be fed into recurrent networks via the cortex. This meant that, for the first time, the brain was able to:
- not only to recognise temporal changes,
- but also to amplify them in a way relevant to attention,
- to learn temporal patterns,
- and recognise them.
This marks a decisive evolutionary step: the emergence of time-processing intelligence.
The inhibitory and time-delayed signal generated by the first and second inversions was already fully capable of producing a difference map during the analogue phase of evolution. As long as the motor input signals were analogue, this two-stage transformation was sufficient to reliably detect movements and highlight temporal changes. It was only with the later digitisation of the signals that two additional inversion stages became necessary to restore the original form of coding and make the signals usable for categorical decisions
15.9The misleading narrative of the direct and indirect pathways in the basal ganglia
The traditional representation of the basal ganglia distinguishes between a ‘direct’ and an ‘indirect’ pathway, which supposedly inhibit or disinhibit the motor thalamus. However, this view describes only the anatomical connections and completely ignores the coding logic and signal transformations that actually take place in the basal ganglia.
From a functional perspective, it is not a matter of inhibition or disinhibition of the motor thalamus, but of something entirely different:
- The inhibitory, doubly inverted and time-delayed analogue signal is utilised in a specific thalamic region to detect movement. This signal is the continuation of the early analogue differentiation that was already established in the double inversion of the striatum and the external globus pallidus.
- The excitatory, quadruple-inverted and digitised signals reach a different thalamic region. These signals do not serve to amplify motor function, but form the excitatory digital signal echoes necessary for recurrent cortical networks (RNNs).
This makes it clear:
The basal ganglia generate two completely different signal streams, which are processed in two different thalamic nuclei — an analogue differential current and a digital recurrent current.
The digital stream is the prerequisite for the cortex to learn and recognise temporal patterns. It is therefore not a motor amplifier, but an RNN driver.
15.10 Example: Recognition of joint angles
The recognition of joint angles demonstrates how signal divergence, extreme value coding and lateral inhibition interact:
- Evolutionary background: Originally, co-activation of motor antagonists → formation of the off-signal.
- Nucleus olivaris: double layers of on/off signals, divergence → minimum-coded joint angle.
- Cerebellar nucleus: Inversion of the minimum → maximum-coded joint angle.
- Thalamus: Lateral inhibition → Sparsely coded joint angle (only one vector position > 0).
Result:
- Precise joint angle detection.
- Energy-efficient, fail-safe, unambiguous.
- All achieved through natural circuits – without artificial neural networks.
Sketch missing!
15.11 Example: Orientation columns in the primary visual cortex V1
Digitisation via divergence modules
- The orientation columns in the primary visual cortex (V1) form the basis for mapping an analogue quantity (orientation angle) onto a finite set of numbers.
- The divergence modules act here like AD converters: they transform continuous input signals into discrete scale values.
- At this point, the term ‘digitisation’ may and should be used – because that is precisely what happens biologically.
Angle vectors instead of continuous angles
- Each eye-dominance column provides a signal vector whose components are orthogonal to one another.
- Only one component is active (corresponding to the detected angle); all others are zero.
- This results in an angle vector that encodes the analogue input value uniquely and sparsely.
- The distance of the inclined line from the centre of the visual field is incorporated into the coordinates of the angle vector. This allows the brain to dispense with the use of bias, which is intended to correct hypersensitivity to small shifts in the coordinate system.
Implications for the architecture
- Our previously described CNN in the pontocerebellum is not rendered redundant by this development, but rather becomes more powerful:
- Previously, it had to recognise angles itself.
- Now it receives the angles already digitised as a vector.
- This allows the Purkinje cells to concentrate directly on pattern recognition – a significant reduction in workload.
Relationship between analogue input and scalar output
- If the orientation angle increases, the extreme value in the angle vector shifts to an adjacent position (e.g. to the right).
- If it decreases, the extreme value shifts in the opposite direction.
- This creates a clear positional encoding: continuous angular changes are represented as discrete shifts in the signal vector.
The transition from analogue image processing to digital processing required that the existing Purkinje cells were able to receive the new input from the orientation columns at all. This necessitated the development of a new projection:
Convolution by Purkinje cells
- Each Purkinje cell receives – without any movement – the analogue output from a total of nine neighbouring retinal ganglion cells and, using its synaptic weights, calculates the scalar product of the input and the weight vector.
- This convolution forms the basis for the integration of signals from small, neighbouring receptive fields.
Signal divergence and new pyramidal cells in V1
- In the visual cortex V1, the analogue signals are distributed across the surface through signal divergence.
- New pyramidal cells integrate the superimposed excitations.
- This results in a sensitivity to periodic signal changes, particularly to angular changes in line elements.
Projection to the cerebellum
- In order for these new pyramidal cells to perform their function, their axons must reach the Purkinje cells, which receive the analogue input from the receptive fields.
- This required the formation of a projection from the cortex via the pons into the mossy fiber system.
- This represents a significant evolutionary step: the direct connection of cortical orientation columns with cerebellar structures.
Later expansion: input to the olivary nucleus
- At a later stage of development, the orientation column pyramidal cells were also able to send their input to the olivary nucleus.
- New climbing fibres ensured that each individual eye-dominance column was represented by a corresponding Purkinje cell.
- This created a precise correspondence between cortical orientation columns and cerebellar processing units.
The emergence of transformer networks in the primate brain
- This architecture – with parallel projections, orthogonal angle vectors and unambiguous Purkinje representation – marks the beginning of the emergence of transformer networks in the primate brain.
- Analogous to technical transformer models, discrete vector positions are used here to efficiently represent and process complex patterns.
Note: Back-transformation in the descending signal pathway
The signals digitised in the secondary brain system must be transformed back into their original analogue signal form in the descending signal pathway from the cortex to the trunk. Only in this way can they fulfil their motor tasks and precisely control the muscles.
This makes it clear that, alongside the divergence structures, there are also convergence structures in the brain that are responsible for converting the digitised data back into analogue control impulses. We will discuss these convergence structures in detail later in this monograph.
Plausibility of signal divergence
Nerve cells are living structures. Their functioning can be impaired or they can even die as a result of a wide variety of factors. It was therefore advantageous for the nervous system to transmit important signals via multiple pathways simultaneously and in parallel. This increased reliability and ensured the stability of information processing.
Over the course of evolution, this developed into the ability to distribute signals divergently across multiple neurons. This signal divergence led – for entirely ordinary, mathematical reasons – to the emergence of more intelligent signal processing. It was the actual starting point for the development of neural, adaptive networks.
Central idea: transition from analogue signals to scaled values
- In the primary brain system, signals were encoded analogously via firing rates.
- With signal divergence came the possibility of mapping these continuous firing rates onto scale values.
- This does not mean ‘digitisation’ in the technical sense, but rather biological scaling: continuous excitation intensities are transformed into clearly distinguishable values.
- This transformation forms the basis for the secondary brain system, in which on-off signal pairs are processed.
15.13 Supplement for AI experts: Orthogonalisation through lateral inhibition
The mapping of the signal strength of analogue signals onto scale values takes place in the secondary brain system through the superposition of on/off signal pairs with a non-linear characteristic curve.
- This superposition leads to the formation of signal vectors whose components are orthogonal to one another following lateral inhibition.
- Orthogonality here means: each component represents an independent signal that is no longer correlated with the others.
- This results in a sparse and unambiguous encoding that avoids redundancy and supports the specialisation of individual neurons.
Comparison with artificial neural networks:
- If an artificial neural network were to perform this transformation, it would require an enormous amount of learning effort and large quantities of electrical energy.
- Nature achieves the same result with a relatively straightforward neural circuit that operates in an energy-efficient manner through lateral inhibition and signal divergence.
- For AI architectures, this suggests that adopting such principles (bus topology, signal-specific inhibition, orthogonalisation) could lead to massive efficiency gains.
15.14 Theorem on the transition from analogue to digital brain systems
The combination of on-off receptors, on-off signal pairs, signal divergence and distance-dependent non-linear signal attenuation led to the emergence of divergence modules in the vertebrate brain, which transform analogue input signals into one-hot signal vectors. This marks the beginning of the secondary digital brain system in vertebrates.
15.15 Summary
At the start of any information processing lies the analogue source quantity: light, sound, pressure or chemical concentration. These continuous signals are detected by receptors and converted into non-linear response functions. Strictly concave or convex transformations thereby generate signal vectors whose extreme value represents the original quantity.
Through inversion, concave vectors are converted into convex ones, ensuring that maximum coding is always present. Subsequently, lateral inhibition ensures that only a single vector component remains active – a sparsely coded vector that reduces the original analogue quantity to a unique scale value.
When several such vectors are superimposed additively, binary patterns with multiple active sites emerge. These digital vectors form the input for the brain’s neural networks. In this way, the transition takes place from the analogue world of sensory stimuli to the digital world of neural processing – a process that not only forms the basis of biological intelligence but also highlights the proximity to artificial intelligence.
15.16 Example: Language acquisition
At this point, reference is made to the internet link https://www.gehirntheorie.de/Kapitel-12.html, which deals extensively with the topic of language acquisition and linguistic ability. The author therefore refrains here from providing a detailed account of the topic ‘language learning’