Signaltheorie der Intelligenz
für das Human Brain Projekt der Europäischen Union
15. Der Übergang zur digitalen Signalverarbeitung im Gehirn
Oft wird betont, das Gehirn unterscheide sich grundlegend von einem Computer. Diese Gegenüberstellung ist jedoch irreführend. Der entscheidende Unterschied scheint auf den ersten Blick in der digitalen Arbeitsweise von Computern zu liegen. Doch auch das Gehirn verarbeitet digitale Signale – und zwar in weit größerem Umfang, als gemeinhin angenommen wird. Nur ein kleiner Teil der neuronalen Aktivität ist tatsächlich analog. Um die weitere Entwicklung biologischer neuronaler Netze zu verstehen, ist es daher notwendig, die Mechanismen der Digitalisierung im Gehirn genauer zu betrachten. Die Digitalisierung neuronaler Signale beruht auf der Differenzierung der Rezeptoren in On-Off-Typen, der nichtlinearen Signaldämpfung auf Axonen und der Überlagerung neuronaler Signale in Divergenzmodulen.
1. Die Differenzierung von Rezeptoren in On-Off-Typen
Der Weg zur Digitalisierung analoger Feuerraten begann mit der funktionellen Differenzierung von Rezeptoren. Aus zunächst ausschließlich „On‑artigen“ Rezeptoren, deren Feuerrate mit zunehmender Signalstärke anstieg, entwickelten sich Rezeptoren, deren Feuerrate mit der Stärke der gemessenen Urgröße abnahm. On‑Rezeptoren zeigen eine streng monoton steigende Antwortfunktion, Off‑Rezeptoren eine streng monoton fallende. Im visuellen System existieren beide Typen sowohl für Helligkeit als auch für Farbwahrnehmung. Ein On‑Signal kann durch Invertierung in ein Off‑Signal überführt werden, indem es ein Dauersignal hemmt. Bei visuellen Rezeptoren entsteht diese Signalinversion durch die Hemmung eines kontinuierlichen Ausgangssignals, das von sogenannten Bandsynapsen erzeugt wird.
Dort, wo die Evolution keine Off-Rezeptoren hervorbrachte, wurden die Off-Signale durch Signalinvertierung im Spinocerebellum gebildet.
Die Grundlage des sekundären Gehirnsystems ist die Trennung von On‑ und Off‑Signalen. Rezeptoren übertragen die Stärke einer Urgröße in eine Feuerrate:
- On‑Signal : Feuerrate steigt mit zunehmender Signalstärke.
- Off‑Signal : Feuerrate sinkt mit zunehmender Signalstärke.
Aus ursprünglich einer Rezeptorenart entstand durch Differenzierung zwei Rezeptorenarten, eine mit dem On-Typ, die andere mit dem Off-Typ. Dies führte auch zur Trennung der Signalwege, so dass Doppelschichten entstanden.
2- On-Off-Signale und die Doppelschichtentheorie nach Malczan
Die Grundlage des sekundären Gehirnsystems ist die Trennung von On‑ und Off‑Signalen. Rezeptoren übertragen die Stärke einer Urgröße in eine Feuerrate:
- On‑Signal : Feuerrate steigt mit zunehmender Signalstärke.
- Off‑Signal : Feuerrate sinkt mit zunehmender Signalstärke.
Diese Dualität führt zur Ausbildung von Doppelschichten im Nervensystem:
- Jede Doppelschicht repräsentiert eine Modalität (z. B. Beugemuskeln vs. Streckmuskeln).
- Im Nucleus olivaris bilden sich solche Doppelschichten für Gelenksignale
- Über die cortikale Projektion entstanden auch cortikale Doppelschichten.
- Im visuellen System findet man diese Doppelschichten z. B. im visuellen Thalamus und (etwas versteckt) im primären visuellen Cortex, wo die Inputsschichten in On-Off-Subschichten angeordnet sind. Auch für die Projektion ins Cerebellum kann eine solche Trennung angenommen werden.
- Mit der Trennung der Rezeptoren in On- und Off-Signale entstand auch eine Trennung der Markersubstanzen in On- und Off-Typen, die zur Organisation der Topologie der Doppelschichten nötig waren.
- Über den Nucleus olivaris wurden diese zwei Markertypen ins Cerebellum übertragen, wo sie zur streifenartigen Kennzeichnung der Cerebellumrinde führten.
Evolutionäre Bedeutung:
- Die Doppelschichtorganisation entstand aus der Koaktivierung motorischer Gegenspieler.
- Experimentelle Befunde (z. B. Zhou et al. 2024) bestätigen die Existenz solcher Markerstrukturen im Cerebellum.
Quellenangabe
Andreas Malczan: Die Doppelschichtentheorie nach Andreas Malczan, Kapitel 17 in Gehirntheorie des Menschen (2024/2025), gehirntheorie.de.
3. Signaldivergenz zur Erhöhung der Ausfallsicherheit
Nervenzellen sind lebende Gebilde. Sie können durch verschiedenste Einflüsse in ihrer Funktionsweise behindert werden oder sogar absterben. Daher war es für das Nervensystem günstig, wichtige Signale auf mehreren Wegen gleichzeitig und parallel zu übertragen. Dies erhöhte die Ausfallsicherheit und sicherte die Stabilität der Informationsverarbeitung. Es entwickelten sich verschiedene Formen der Signaldivergenz.
Formen der Signaldivergenz:
- Lineare Divergenz: Ein Axon verzweigt sich entlang seiner Länge. Andere Neuronen zapfen diese Verzweigungen an, sodass sich die Erregung auf mehrere Outputneuronen verteilt.
- Flächenhafte Divergenz: Axonbüschel oder Interneuronen verteilen die Erregung in der Ebene. Viele Outputneuronen greifen auf diese verteilten Signale zu, wodurch sich der Input über eine Fläche verteilt.
- Räumliche Divergenz: Die Erregung breitet sich dreidimensional aus und wird von verschiedenen Neuronen in unterschiedlichen Regionen aufgenommen.
Eine Folge der Signaldivergenz war die Signaldämpfung in Abhängigkeit von der Weglänge der Signale.
4. Signaldämpfung und Weglänge
Signale werden auf Axonen übertragen. Ständig offene Ionenkanäle entlang der Axone führen zur Veränderung der Ionenkonzentration im Axon, wenn Ionen einströmen oder das Axon verlassen. Dadurch nimmt die Erregung mit zunehmendem Abstand ab, sodass nur ein Teil der ursprünglichen neuronalen Aktivität im Outputneuron ankommt.
Die Abnahme der Erregung als Funktion des Abstandes wird hier als Übertragungsfunktion bezeichnet:
- Entlang eines einzelnen Axons ist sie exponentiell und strikt konkav, entsprechend der klassischen Kabelgleichung (entwickelt und gemessen u. a. von Wilfrid Rall in den 1960er Jahren):
![]()
- In der Ebene ist die Dämpfung stärker, da der Flächeninhalt quadratisch mit dem Durchmesser steigt; die Übertragungsfunktion ist hier strikt konvex:
![]()
- Bei der räumlichen Ausbreitung ist die Übertragungsfuktion prinzipiell identisch mit der Ausbreitung in der Ebene, denn man kann sich vorstellen, alle Axone so zurechtzubiegen, dass sie wieder in einer Ebene liegen.
5. Signalüberlagerung in Divergenzmodulen
In Divergenzmodulen überlagern sich neuronale Erregungen mehrerer Signale additiv.
- Treffen mehrere Erregungen aus unterschiedlichen Orten zusammen, überlagern sich ihre Übertragungsfunktionen.
- Diese Überlagerungen besitzen stets einen globalen Extremwert:
- Lineare Divergenzmodule → Signalminimum.
- Flächenhafte Divergenzmodule → Signalmaximum.
- Räumliche Divergenzmodule → Signalmaximum.
- Das Signalminimum der linearen Module wird im Spinocerebellum invertiert und in ein maximumcodiertes Signal überführt. Das Spinocerebellum wirkt hier als Invertierungsmodul.
- Der globale Extremwert verschlüsselt eineindeutig den Wert der Urgröße des Rezeptors über das Signalstärkeverhältnis des On-Off-Signalpaares.
Beispiel für ein lineares Divergenzmodul
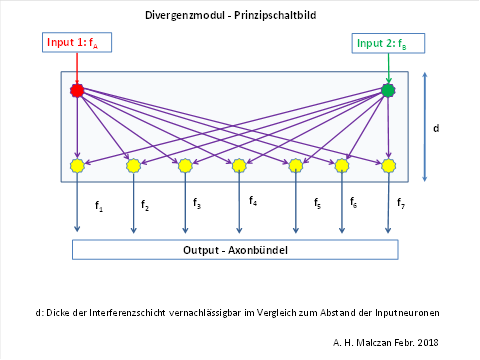
In einem ebenen Divergenzmodul sind die Inputneuronen z. B. an den Ecken eines Quadrates angeordnet, während die Outputneuronen innerhalb dieser Fläche ungefähr gleichmäßig verteilt sind. Jedes Outputneuron ist mit jedem Inputneuron synaptisch verbunden.
6. Laterale Hemmung und Gaußsche Kodierung
Die Entstehung einer maximumcodierten Übertragungsfunktion in Divergenzmodulen bzw. im nachfolgenden Invertierungsmodul ermöglicht den Gehirn die Transformation analoger Signale in eine Gaußsche Kodierung (Gaussian Encoding). Hierbei wird eine stetige Feuerrate auf eine diskrete Skala abgebildet, indem das Neuron mit der maximalen Erregung durch laterale Hemmung die Erregung der übrigen Neuronen teilweise unterdruckt. So bleibt ein glockenförmiger Erregungsverlauf übrig, dessen Maximum die Signalstärke repräsentiert.
- Das Neuron, dessen Membranpotential durch zeitliche und räumliche Summation der Inputs das Maximum darstellt, erreicht als erstes die Auslöseschwelle für ein Aktionspotential.
- Dieses Neuron feuert als erstes und aktiviert unmittelbar die hemmenden Interneuronen.
- Die hemmenden Interneuronen unterdrücken die nur teilweise angereicherte Erregung aller Konkurrenzneuronen.
Damit greift das bekannte Konkurrenzprinzip der Neuronen:
„The winner takes all.“
Folgen für die Signalverarbeitung:
- Aus dem maximumkodierten Signalvektor im efferenten Signalweg wird ein spärlich kodierter Vektor.
- Nur wenige Vektorpositionen besitzt einen Wert größer als Null. Auf allen anderen Positionen wird die Erregung durch eine Art Gaußkurve begrenzt. Vorteil: Stetigkeit der Abbildung
- Dies entspricht der klassischen Gaußschen Kodierung.
- Sollte das aktuell meisterregte Neuron ausfallen, übernimmt ein Nachbarneuron das Signalmaximum.
- Dadurch werden neuronale Schaltungen extrem ausfallsicher und zugleich energieeffizient.
Die Übertragung analogen Feuerraten in einen Vektor mit Gaußsche Kodierung stellt die Digitalisierungsmethode des Nervensystems der Wirbeltiere dar. Diese Vektoren der analogen Feuerrate eines Rezeptors sind untereinander fast orthogonal und bestens geeignet für die Signalauswertung in verschiedensten Neuronalen Netzen wie KNNs, CNNs oder RNNs. Sie sind auch die Signalgrundlage für die Entstehung der höchsten Netzklasse, der Transformer.
7. Die digitale Evolution der Basalganglien
Die frühen Wirbeltiergehirne arbeiteten ausschließlich mit analogen Signalen. Die Differenzbildung zur Bewegungserkennung entstand im Striatum, das ursprünglich nur aus Striosomen bestand. In diesen befanden sich GABAerge Projektionsneurone, die vom dopaminergen Mittelwertkern motorische Ausgangssignale erhielten. Zwei unterschiedlich lange Signalwege – einer erregend, einer hemmend – ermöglichten die Auslöschung zeitkonstanter Signale und die Verstärkung zeitlicher Veränderungen. Dies war die erste analoge Differenzschaltung des Nervensystems.
Mit zunehmender Komplexität der Motorik reichte die analoge Differenzbildung nicht mehr aus. Motorische Entscheidungen mussten nicht nur graduell, sondern kategorial getroffen werden. Dies führte zur Digitalisierung der Signale: analoge Eingangswerte wurden in diskrete, one‑hot‑ähnliche Signalvektoren überführt. Um diese neuen digitalen Repräsentationen verarbeiten zu können, wurde das Striatum erweitert. Neben den Striosomen entstanden die Matrix‑Zonen, die die Grundlage der heutigen Go/No‑Go‑Schaltung bilden. Damit wandelten sich die Basalganglien von einem analogen Differenzdetektor zu einem digitalen Entscheidungsmodul, das kategoriale motorische Optionen auswählt und verstärkungsbasiert optimiert.
8. Erregende Rekurrenz: Die vierfache Signalinversion als Voraussetzung für RNN‑Funktionalität 8.
Rekurrente neuronale Netze benötigen erregende Signalechos, um zeitliche Muster zu stabilisieren, zu integrieren und wiederzuerkennen. Die frühen Basalganglien erzeugten jedoch ausschließlich hemmende Echos. Damit diese Signale in rekurrente kortikale Netzwerke eingespeist werden konnten, musste das Gehirn eine Lösung finden, die hemmenden, verzögerten Signale wieder in erregende, maximumcodierte Signale zurückzuverwandeln.
Dies geschah durch eine evolutionär entstandene vierfache Signalinversion, die sich über mehrere Kerngebiete erstreckt:
Erste Inversion – Striatum
Hemmung eines Dauersignals erzeugt die notwendige Monotonieumkehr, aber führt zu einer minimumcodierten Repräsentation. Gabaerge Projektionsneuronen mir großen dendritischen Feldern empfingen eine Art Dauererregung von den hindurchziehenden cortikalen Parymidenzellen. Aus der Substantia nigra pars compacta empfingen diese eine Signalkopie der Cortexsignale, welche die Striatumneuronen hemmte. Dies war die erste Inversion der cortikalen Signale.
Zweite Inversion – Globus pallidus externa
Das ursprünglich einheitliche pallidale System spaltete sich funktional auf. Der GPe invertiert das minimumcodierte Signal erneut, um die ursprüngliche Codierungsform wiederherzustellen, jedoch weiterhin hemmend und verzögert. Sie wird für die Bewegungsanalyse durch eine Fifferenzschaltung benötigt. Hierbei hemmt das zeitverzögerte Signal das zugehörige aktuelle Signal im Thalamus. Das Dauersignal zur Inversion liefert der Nucleus subthalamicus.
Dritte Inversion – Globus pallidus interna
Der GPi übernimmt eine weitere Inversion, auch hier liefert der Nucvleus subthalamicus das nötige Dauersignal für die Inversion.
Vierte Inversion – Thalamus
Die letzte Inversion erfolgt an einem erregenden Outputneuron im thalamischen Mittelwertkern. Erst hier wird das Signal wieder erregend, maximumcodiert und bleibt gleichzeitig zeitverzögert.
Nach dieser vierstufigen Transformation können die Signale über den Cortex in rekurrente Netzwerke eingespeist werden. Damit war das Gehirn erstmals in der Lage:
- zeitliche Veränderungen nicht nur zu erkennen,
- sondern auch aufmerksamkeitsrelevant zu verstärken,
- zeitliche Muster zu erlernen ,
- und wiederzuerkennen.
Dies markiert einen entscheidenden evolutionären Schritt: Die Entstehung zeitverarbeitender Intelligenz.
Das durch die erste und zweite Inversion erzeugte hemmende und zeitverzögerte Signal war in der analogen Phase der Evolution bereits vollständig geeignet, eine Differenzabbildung zu erzeugen. Solange die motorischen Eingangssignale analog waren, reichte diese zweistufige Transformation aus, um Bewegungen zuverlässig zu erkennen und zeitliche Veränderungen hervorzuheben. Erst mit der späteren Digitalisierung der Signale wurden zwei zusätzliche Inversionsstufen notwendig, um die ursprüngliche Codierungsform wiederherzustellen und die Signale für kategoriale Entscheidungen nutzbar zu machen
9. Die irreführende Erzählung vom direkten und indirektem Weg in den Basalganglien
Die traditionelle Darstellung der Basalganglien unterscheidet zwischen einem „direkten“ und einem „indirekten“ Weg, die angeblich den motorischen Thalamus hemmen oder enthemmen. Diese Sichtweise beschreibt jedoch ausschließlich die anatomische Verschaltung und ignoriert vollständig die Codierungslogik und die Signaltransformationen, die in den Basalganglien tatsächlich stattfinden.
Funktional betrachtet geht es nicht um Hemmung oder Enthemmung des motorischen Thalamus, sondern um etwas völlig anderes:
- Das hemmende, doppelt invertierte und zeitverzögerte analoge Signal wird in einem spezifischen thalamischen Gebiet genutzt, um Bewegung zu erkennen. Dieses Signal ist die Fortsetzung der frühen analogen Differenzbildung, die bereits in der zweifachen Inversion des Striatums und des Globus pallidus externa angelegt war.
- Die erregenden, vierfach invertierten und digitalisierten Signale erreichen ein anderes thalamisches Gebiet. Diese Signale dienen nicht der Verstärkung der Motorik, sondern bilden die erregenden digitalen Signalechos, die für rekurrente kortikale Netzwerke (RNNs) notwendig sind.
Damit wird klar:
Die Basalganglien erzeugen zwei völlig unterschiedliche Signalströme, die in zwei verschiedenen Thalamuskernen verarbeitet werden — einen analogen Differenzstrom und einen digitalen Rekurrenzstrom.
Der digitale Strom ist die Voraussetzung dafür, dass der Cortex zeitliche Muster erlernen und wiedererkennen kann. Er ist also kein Motorverstärker, sondern ein RNN‑Treiber.
10. Beispiel: Erkennung von Gelenkwinkeln
Die Erkennung von Gelenkwinkeln zeigt, wie Signaldivergenz, Extremwertkodierung und laterale Hemmung zusammenwirken:
- Evolutionärer Hintergrund : Ursprünglich Koaktivierung motorischer Gegenspieler → Bildung des Off‑Signals.
- Nucleus olivaris : Doppelschichten von On/Off‑Signalen, Divergenz → Minimumkodierter Gelenkwinkel.
- Kleinhirnkern : Inversion des Minimums → Maximumkodierter Gelenkwinkel.
- Thalamus : Laterale Hemmung → Spärlich kodierter Gelenkwinkel (nur eine Vektorposition > 0).
Ergebnis:
- Präzise Gelenkwinkelerkennung.
- Energieeffizient, ausfallsicher, eindeutig.
- Alles durch natürliche Schaltungen – ohne künstliche neuronale Netze.
Skizze fehlt!
11. Beispiel: Orientierungssäulen im primären visuellen Cortex V1
Digitalisierung durch Divergenzmodule
- Die Orientierungssäulen im primären visuellen Cortex (V1) bilden die Grundlage für die Abbildung einer analogen Größe (Orientierungswinkel) auf eine endliche Zahlenmenge.
- Die Divergenzmodule wirken hier wie AD‑Wandler: sie transformieren kontinuierliche Eingangssignale in diskrete Skalenwerte.
- An dieser Stelle darf und soll der Begriff Digitalisierung verwendet werden – denn genau das geschieht biologisch.
Winkelvektoren statt kontinuierlicher Winkel
- Jede Augendominanzsäule liefert einen Signalvektor, dessen Komponenten orthogonal zueinander sind.
- Nur eine Komponente ist aktiv (entspricht dem erkannten Winkel), alle übrigen sind Null.
- Damit entsteht ein Winkelvektor, der die analoge Eingangsgröße eindeutig und spärlich kodiert.
- Der Abstand der geneigten Linie vom Bildmittelpunkt des Sichtfeldes geht in die Koordinaten des Winkelvektors ein. Dadurch kann im Gehirn auf die Verwendung des Bias verzichtet werden, der eine Überempfindlichkeit auf kleine Verschiebungen des Koordinatensystems beheben soll.
Folgen für die Architektur
-
Unser
bereits beschriebenes CNN im Pontocerebellum wird durch diese Entwicklung
nicht überflüssig, sondern leistungsfähiger:
- Früher musste es Winkel selbst erkennen.
- Nun erhält es die Winkel bereits fertig digitalisiert als Vektor.
- Die Purkinjezellen können sich dadurch direkt auf die Mustererkennung konzentrieren – eine deutliche Arbeitserleichterung.
Beziehung zwischen analogem Input und Skalenoutput
- Vergrößert sich der Orientierungswinkel, wandert der Extremwert im Winkelvektor auf eine benachbarte Position (z. B. nach rechts).
- Verkleinert er sich, wandert der Extremwert in die entgegengesetzte Richtung.
- Damit entsteht eine klare Positionskodierung: kontinuierliche Winkeländerungen werden als diskrete Verschiebungen im Signalvektor abgebildet.
Der Übergang von der analogen Bildverarbeitung zu digitaler Verarbeitung erforderte, dass die bisherigen Purkinjezellen den neuen Input der Orientierungssäulen überhaupt empfangen konnten. Dazu war die Herausbildung einer neuen Projektion nötig:
Faltung durch Purkinjezellen
- Jede Purkinjezelle empfängt – ohne jegliche Bewegung – den analogen Output von insgesamt neun benachbarten retinalen Ganglienzellen und bildet mittels ihrer synaptischen Gewichte das Skalarprodukt aus Input und Gewichtsvektor.
- Diese Faltung bildet die Grundlage für die Integration von Signalen aus kleinen, benachbarten rezeptiven Feldern.
Signaldivergenz und neue Pyramidenzellen in V1
- Im visuellen Cortex V1 werden die analogen Signale durch Signaldivergenz in der Fläche verteilt.
- Neue Pyramidenzellen integrieren die sich überlagernden Erregungen.
- Dadurch entsteht eine Empfindlichkeit auf periodische Signaländerungen, insbesondere auf Winkeländerungen von Linienelementen.
Projektion ins Kleinhirn
- Damit diese neuen Pyramidenzellen ihre Funktion entfalten können, müssen ihre Axone die Purkinjezellen erreichen, die den analogen Input aus den rezeptiven Feldern empfangen.
- Dazu war die Ausbildung einer Projektion vom Cortex über die Pons in das Moosfasersystem nötig.
- Dies stellt einen bedeutenden Evolutionsschritt dar: die direkte Verbindung von kortikalen Orientierungssäulen mit cerebellaren Strukturen.
Spätere Erweiterung: Input zum Nucleus olivaris
- In einer späteren Entwicklungsstufe konnten die Orientierungssäulen‑Pyramidenzellen ihren Input auch zum Nucleus olivaris senden.
- Neue Kletterfasern sorgten dafür, dass jede einzelne Augendominanzsäule durch eine zugehörige Purkinjezelle repräsentiert wurde.
- Damit entstand eine präzise Zuordnung zwischen kortikalen Orientierungssäulen und cerebellaren Verarbeitungseinheiten.
Beginn der Transformer‑Netzwerke im Gehirn der Primaten
- Diese Architektur – mit parallelen Projektionen, orthogonalen Winkelvektoren und eindeutiger Purkinje‑Repräsentation – markiert den Beginn der Herausbildung von Transformer‑Netzwerken im Gehirn der Primaten.
- Analog zu technischen Transformer‑Modellen werden hier diskrete Vektorpositionen genutzt, um komplexe Muster effizient zu repräsentieren und weiterzuverarbeiten.
12. Hinweis: Rücktransformation im absteigenden Signalweg
Die im sekundären Gehirnsystem digitalisierten Signale müssen im absteigenden Signalweg vom Cortex zum Rumpf wieder in ihre ursprüngliche analoge Signalform zurücktransformiert werden. Nur so können sie ihre motorischen Aufgaben erfüllen und die Muskeln präzise ansteuern.
Damit wird deutlich: Parallel zu den Divergenzstrukturen existieren im Gehirn auch Konvergenzstrukturen, die für die Rückführung der digitalisierten Daten in analoge Steuerimpulse verantwortlich sind. Auf diese Konvergenzstrukturen gehen wir im weiteren Verlauf der Monografie noch ausführlich ein.
Plausibilität der Signaldivergenz
Nervenzellen sind lebende Gebilde. Sie können durch verschiedenste Einflüsse in ihrer Funktionsweise behindert werden oder sogar absterben. Daher war es für das Nervensystem günstig, wichtige Signale auf mehreren Wegen gleichzeitig und parallel zu übertragen. Dies erhöhte die Ausfallsicherheit und sicherte die Stabilität der Informationsverarbeitung.
Im Verlauf der Evolution entwickelte sich daraus die Fähigkeit, Signale divergent auf mehrere Neuronen zu verteilen. Diese Signaldivergenz führte – aus ganz gewöhnlichen, mathematischen Gründen – zur Herausbildung einer intelligenteren Signalverarbeitung. Sie war der eigentliche Startschuss für die Entwicklung neuronaler, lernfähiger Netze.
Zentrale Idee: Übergang von analogen Signalen zu Skalenwerten
- Im primären Gehirnsystem wurden Signale durch Feuerraten analog codiert.
- Mit der Signaldivergenz entstand die Möglichkeit, diese kontinuierlichen Feuerraten auf Skalenwerte abzubilden.
- Dies bedeutet keine „Digitalisierung“ im technischen Sinn, sondern eine biologische Skalierung: kontinuierliche Erregungsstärken werden in klar unterscheidbare Werte transformiert.
- Diese Transformation bildet die Grundlage für das sekundäre Gehirnsystem, in dem On‑Off‑Signalpaare verarbeitet werden.
13. Ergänzung für KI-Experten: Orthogonalisierung durch laterale Hemmung
Die Abbildung der Signalstärke von analogen Signalen auf Skalenwerte erfolgt im sekundären Gehirnsystem durch die Signalüberlagerung von On/Off‑Signalpaaren mit nichtlinearer Kennlinie.
- Diese Überlagerung führt zur Bildung von Signalvektoren, deren Komponenten nach einer lateralen Hemmung orthogonal zueinander sind.
- Orthogonalität bedeutet hier: Jede Komponente repräsentiert ein unabhängiges Signal, das nicht mehr mit den anderen korreliert ist.
- Damit entsteht eine sparsame und eindeutige Kodierung, die Redundanz vermeidet und die Spezialisierung einzelner Neuronen unterstützt.
Vergleich mit künstlichen neuronalen Netzen:
- Würde ein künstliches neuronales Netz diese Transformation durchführen, wären dafür ein enormer Lernaufwand und große Mengen an elektrischer Energie erforderlich.
- Die Natur erreicht dasselbe Resultat mit einer relativ unkomplizierten neuronalen Schaltung, die durch laterale Hemmung und Signaldivergenz energiesparend arbeitet.
- Für KI‑Architekturen ist dies ein Hinweis, dass die Übernahme solcher Prinzipien (Bustopologie, signalspezifische Hemmung, Orthogonalisierung) zu massiven Effizienzgewinnen führen könnte.
14. Theorem des Übergangs von analogen zum digitalen Gehirnsystem
Die Kombination aus On‑Off‑Rezeptoren, On‑Off‑Signalpaaren, Signaldivergenz und abstandsabhängiger nichtlinearer Signaldämpfung führte im Wirbeltiergehirn zur Entstehung von Divergenzmodulen, welche analoge Eingangssignale in One‑Hot‑Signalvektoren transformieren. Dies markiert den Beginn des sekundären digitalen Gehirnsystems der Wirbeltiere.
15. Zusammenfassung
Am Beginn jeder Informationsverarbeitung steht die analoge Urgröße: Licht, Schall, Druck oder chemische Konzentration. Diese kontinuierlichen Signale werden durch Rezeptoren aufgenommen und in nichtlineare Antwortfunktionen überführt. Strikt konkave oder konvexe Transformationen erzeugen dabei Signalvektoren, deren Extremwert die ursprüngliche Größe repräsentiert.
Durch Inversion werden konkave Vektoren in konvexe überführt, sodass stets eine Maximumkodierung vorliegt. Anschließend sorgt laterale Hemmung dafür, dass nur eine einzige Vektorkomponente aktiv bleibt – ein spärlich kodierter Vektor, der die analoge Urgröße auf einen eindeutigen Skalenwert reduziert.
Werden mehrere solcher Vektoren additiv überlagert, entstehen binäre Muster mit mehreren aktiven Stellen. Diese digitalen Vektoren bilden die Eingabe für die neuronalen Netze des Gehirns. Auf diese Weise vollzieht sich der Übergang von der analogen Welt der Sinnesreize zur digitalen Welt der neuronalen Verarbeitung – ein Prozess, der nicht nur die Grundlage biologischer Intelligenz bildet, sondern zugleich die Nähe zur künstlichen Intelligenz verdeutlicht.
16. Beispiel: Sprache erlernen
An dieser Stelle sei auf den Internet-Link https://www.gehirntheorie.de/Kapitel-12.html verwiesen, der das Thema Spracherwerb und Sprachvermögen ausführlich behandelt. Daher verzichtet der Autor hier auf die ausführliche Darstellung zum Thema „Sprache erlernen“
Andreas Heinrich Malczan