Human brain theory
for the European Union’s Human Brain Project
ISBN 978-3-00-068559-0
14.5 Movement control through centre of gravity modules
The spine allows movement in all directions. The person's body can be bent forward or backward, as well as to the left or to the right. One can also let the upper body circle.
A similar mobility can be observed in fish, but also in snakes, as well as in worms.
Such mobility in vertebrates requires joints that have several degrees of freedom. As a model, let us assume that two pairs of muscles (in the minimum case) would be sufficient to enable such movements. One pair of muscles enables movement in one plane, the second pair of muscles in a second, inclined or perpendicular plane.
Four muscles require four control signals. Two muscles each form a pair of antagonistic muscles that are excited inversely to each other. For them, we need a pair of neuronal signals that are inverse to each other.
For the second pair of muscles, we also need such a pair of mutually inverse signals.
Who can provide the appropriate signals?
To answer this question, we turn to the purpose of muscle movement. Joints with more than one degree of freedom allow rotational movements. The trunk can be rotated, as can the head. But also the arms and the legs can be rotated in a circle if necessary. Such a movement is usually controlled by the eyes. If you stop the movement, you achieve a bending in a certain direction.
We are trying to use the signals from the eyes to control the movement of an outstretched arm that is supposed to grasp an apple, for example. As a model, we are investigating the movement in a human being, as their brain structures are well researched.
The output signals come from the retina. We decide to use the right eye as an example. Its signals arrive in the sensory thalamus via the optic nerve and reach the primary visual cortex, but also a structure known as the tectum opticum. In vertebrates, this is also known as the superior colliculus and is present bilaterally. However, only the signals from the magnocellular retinal ganglion cells with particularly large receptive fields (presumably) arrive there. In this respect, a mean value image of the retina will be present in the tectum opticum.
The axons of the visual projection neurons into the tectum may end in a surface that represents an average image of the retina. In higher vertebrates - due to the formation of different types of visual receptors - there may well be several such input surfaces, for example one for light/dark, possibly even for different colours. In this case, they are stacked in layers. We decide on the tectal input surface dark-on/light-off. It reacts to dark objects in front of a light background.
There, the axons of the visual mean neurons of this visual modality end in an almost circular area, which we can imagine as the mean image of the retina. We assume that during the transmission of the visual signals into the tectum opticum, the fidelity of the image is maintained, i.e. the image is retinotopic.
In the Tectum opticum we imagine a virtual coordinate system consisting of a u-axis and a v-axis. The u-axis runs horizontally through the centre of the field of vision, the v-axis vertically through the centre of the field of vision. If we look straight ahead, we can assign the coordinates u = 0 and v = 0 to the point of view. We decide on the variables u and v because we will later need a second coordinate system with an x-axis and a y-axis.
If we imagine a circle with a certain radius in the plane of the visual field, the centre of which runs through the centre of the visual field, this circle is also depicted as a circle in the projection surface via the visual projection from the retina to the tectum opticum.
Now let the visual background be a white wall on which a small black circle (for example a small black ball) rotates exactly on a circular line around the centre of the field of vision. This could be achieved, for example, by visually simulating this event in the computer and projecting it onto the wall via a projector.
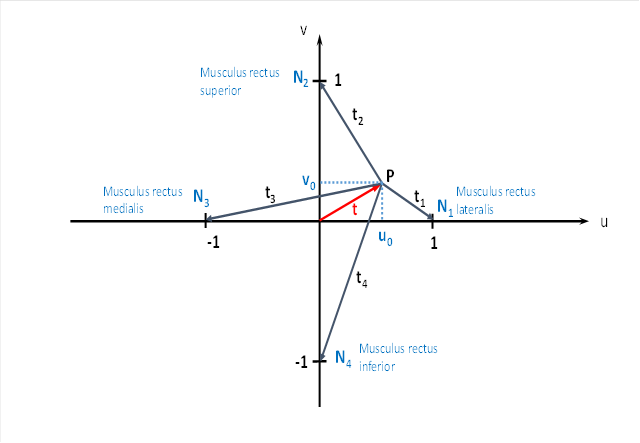
The image of the rotating black ball in the tectum can now be described by two quantities. It is located at the point P. Firstly, it has a distance t from the centre. Secondly, the connecting line from P to the centre has an angle ω with the u-axis. Alternatix one can also simply give the u-coordinate and the v-coordinate of the apple centre. Both are related to the distance and the angle via the Pythagorean theorem and via trigonometric functions.
The following applies
![]()
![]()
We call the angle ω the phase angle and t the radius vector in the primal size diagram.
We recall that the tectum is a layered structure, in the upper layers the retinal output ends separately by modality: Light-On/Dark-Off, Dark-On/Light-Off, Red-On/Green-Off, Green-On-Red-Off, Blue-On/Yellow-Off.
We focus on the horizontal and circular layer of the retinal signals dark-on/light-off, which responds best to black objects against a white background. If the background had a different colour and the ball a different, complementary colour to it, the mathematical considerations would be identical, but would refer to a different visual input layer of the tectum.
We therefore assume that the black ball at the point on the retina where its image is located excites the receptors to the maximum. All other receptors of the dark-on/light-off type are completely inactive.
A retinal ganglion cell in the centre of the image may pick up the retinal excitation of the eye with its larger dendrite tree and send it to the tectum. The other retinal ganglion cells are inactive.
Therefore, visual excitation occurs at location P of the tectum, and all other input neurons in this tectum layer receive no signal. (We are looking at one-eyed vision here and therefore only one tectum out of two).
Now the excitation will spread from point P via interneurons to the four neurons that we have labelled N1 , N2 , N3 and N4 in the figure. Each neuron receives an excitation share that depends on the distance to point P. We have labelled these four distances t1 , t , t23 and t .4
Why are these neurons in the places where we have drawn them? The drawn neurons send their signals to four eye muscles and four neck muscle groups. We will name more projection targets later.
We refer here to the right eye.
The neuron N1 projects to the eye muscle Musculus rectus lateralis, which turns the gaze outwards. At the same time, this neuron can activate the neck muscle group that turns the head to the right.
The neuron N2 sends its axon from the tectum opticum upwards to the eye muscle called musculus rectus superior. When contracted, this muscle causes the eye to move upwards, i.e. the gaze moves upwards in the direction of the u-axis. We want to neglect a small inward deflection of the eyeball here, as we are only concerned with the principle.
Downwards, the neuron N2 projects to those neck muscles that cause the head to be lifted upwards.
The neuron N3 activates the eye muscle Musculus rectus medialis, which turns the gaze inwards (if we take the right eye as a basis). Likewise, this neuron activates those neck muscles that cause the head to turn to the left.
The neuron N4 of the tectum excites, on the one hand, the rectus inferior muscle at the eye, which causes a downward eye movement so that the gaze is lowered. At the same time, the neck muscles are excited, which lower the head.
If we had chosen the left eye, left and right would have to be swapped!
But where do the eye muscles get their excitation from? It is quite simple. The excitations of the eye muscles are caused by the eye. The visual mean signals from the retina project into the visual input layer of the tectum and excite the visual input neurons there, which are distributed in this area in the same way as the mean neurons of the eye. In this way, the fidelity of the image is maintained, the image is retinotopic.
The excitation spreads from the excitation centre P to the four output neurons of the tectum via interneurons. During the propagation of the excitation, a signal attenuation occurs that increases exponentially with the square of the distance. We want to calculate the excitation of the four output neurons.
As input excitation, we take a firing rate fp , which arrives at point P of the tectum coming from the retina and may represent the image of the black ball. From point p, the excitation fp spreads to the four output neurons.
Since the excitation reaching the neurons N1 , N2 , N3 and N4 depends on the square of the distance to the point P, we first calculate the squares.
![]()
![]()
![]()
![]()
If we denote by fp the firing rate that will arrive in the tectum opticum at point P coming from the retina, we can calculate the firing rates f1 , f2 , f3 and f4 that will arrive in the neurons N , N , N123 and N . 4
The formulas for this are:
![]()
![]()
![]()
![]()
The common factor
![]()
We bracket out and get
![]()
![]()
![]()
We note that the variable fm contains only the variables u0 and v0 or the vector t , the variables x or y do not occur here. The quantity λ is the longitudinal constant of the neurons.
It is easy to see that all four fire rates are periodic functions of the angle ω and have the period 2π.
So if the black ball in front of a white background rotates around the centre of the field of vision, the gaze follows this object and the direction of gaze also rotates around the centre, because the eye muscle signals are periodic functions.
With this, we calculated the firing rates of the neurons N1 to N4 , which reach the associated eye muscles headward on the axons of these neurons and which also project to the four neck muscle groups for head movement.
We hypothesise that these neuronal excitations propagate up and down the axons from the tectum opticum without changing the firing rate and arrive in different target neurons.
As long as the visual object is outside the centre of the field of vision, the signals generated by the focal module for the motoneurons of the eye muscles are different, so that a movement of the eye takes place. Only when the object reaches the centre of the image does the eye movement end. Then the signals for the motoneurons of the eye muscles all have the same firing rate.
So when a black fly walks back and forth on a white wall, it is not controlling the eye muscles with imaginary forces. It is the visual image of the fly on the retina that excites the light-sensitive receptors. This excitation enters the tectum opticum and spreads to its edge, where the projection neurons to the motoneurons of the eye muscles are located. Due to the different distances of the fly image to the tectum edge, the signal attenuation of these signals is different. Therefore, the eye muscles receive different contraction commands as long as the fly is not centrally located in the field of view.
In a similar way, a vestibular centre of gravity module in the torus semicircularis controls the muscles of the body so that we remain in a stable position as much as possible and do not fall down. Here, the control is more complex because many joints are connected in series and joint angles must be offset against each other. Layered centre of gravity modules are perfectly capable of doing this. However, the mathematical proof is somewhat more complicated and is not given here.
Monografie von Dr. rer. nat. Andreas Heinrich Malczan