Human brain theory
for the European Union’s Human Brain Project
ISBN 978-3-00-068559-0
14.6 Sensorimotor protocol modules
In the previous chapter we had analysed the movement control through centre of gravity modules. Now we go one step further. We know that the output of the focal grid in the case of the tectum controls the motoneurons of the eye muscles. But the eye muscles also produce an output that informs about their muscle tension. Tendon organs are excited by muscle tension and produce a firing rate that increases as muscle tension increases.
Here we can compare the way the tendon organs work in terms of physical principle with spiral springs, whose increase in length is a measure of the muscle tension.
For the eye muscle, we assume for simplicity that the tension of its tendon organ is proportional to the angle through which the eye moves.
To do this, we imagine the field of vision with a coordinate system. The horizontal axis u stands for the lateral movement of the gaze, the vertical axis v refers to the vertical eye movements. Often both movement components are present at the same time. If the eye then fixes the point P, we can use the quantities u0 and v0 as a measure of the tensile stress of the tendon organs of the associated four eye muscles, which are entered in the coordinate system with their names. These tensile tensions are translated into neuronal firing rates.
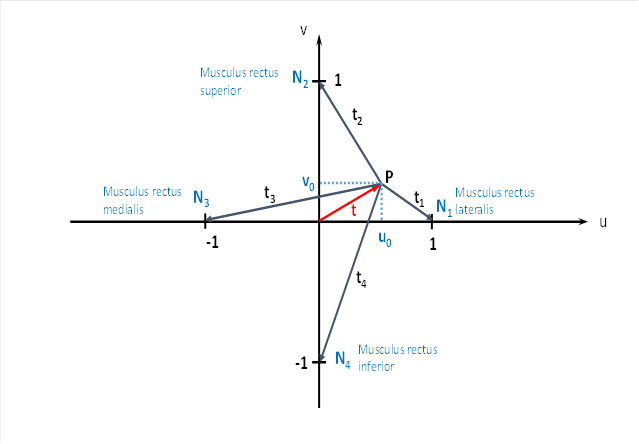
The quantity u0 is responsible for the lateral movement of the eye to the left and right. For a movement to the right, u0 becomes positive, for the opposite direction negative.
The size v0 is responsible for raising and lowering the gaze. Positive values of v0 mean an upward eye movement, negative a downward movement.
If u0 = 0 and v0 = 0 is true, we look straight ahead.
The following applies for the variables involved u0 , v0 , t and ω here
![]()
![]()
![]()
We call the angle ω the phase angle and t the radius vector in the primal size diagram. The phase angle is the angle between the positive x-axis and the radius vector t.
The tendon organs of the eye muscles transfer the tensile stress into a rate of fire. We do not know the concrete transfer function, but we do know that the rate of fire must increase with increasing tensile stress. We also suspect an exponential transfer characteristic.
Let an average firing rate fm be the initial value. Let it be present for all four eye muscles when the gaze is directed straight ahead, i.e. u0 = 0 and v0 = 0.
For the fire rates of the four eye muscles we choose the following (theoretical) approach:
![]()
![]()
![]()
![]()
A comparison with the firing rates of the centre of gravity module shows that the firing rates are just as large as in the case where an object is located at point P and the gaze is directed straight ahead, as is the case in the tectal centre of gravity module. We assume a certain compatibility here.
So if the vertical or horizontal angle of attack of the eye changes, the firing rate changes according to the above formulas. These are only to be understood as example formulas until neurologists and optometrists have actually measured the tension of the tendon organs of the eye muscles. In any case, the rate of fire increases with increasing angle of deflection; the rate of fire functions are strictly monotonically increasing. Anyone is free to follow the calculations with a different theoretical approach, but the result will in principle remain the same.
The tendon organs project headward to the cortical floor in the cord ladder nervous system and in the neural tube. In the chapter on the motor divergence module with lateral signal transmission, it was shown that in the cortical projection field of these signals, excitation maxima move around a centre as the joint angles change.
We want to discuss this using the eye as an example.
N1 to N4 are the cortical neurons that receive the signals from the tendon organs of the four eye muscles.
We graphically represent the cortical area where the eye muscle signals from the tendon organs arrive in a coordinate system.
The excitations f1 to f4 arrive at points N1 to N .4
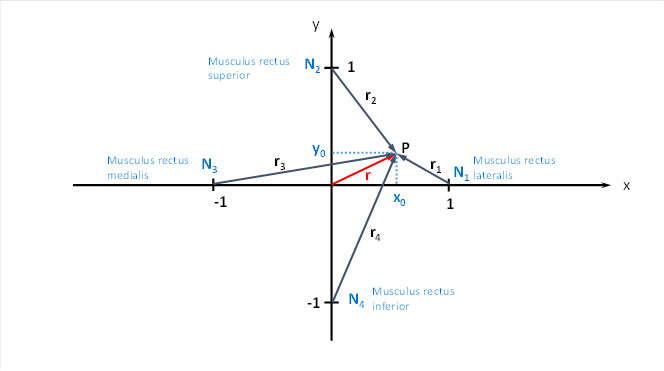
This time the output of the four tendon organs of the eye muscles arrives in the four neurons N1 to N4 . Like any neuronal excitation, it can propagate in the cortical area, for which it uses interneurons. We have always assumed in this monograph that the transfer function for neuronal excitation is generally a strictly concave function over a relatively large area. If four such excitations from the neurons N1 and N4 overlap, the sum again produces a strictly concave function with a global excitation maximum. If the direction of the eye's gaze changes, the location of the maximum excitation also changes. If the eyes follow a rotational movement, the vector t in the above representation also rotates. Each angle constellation leads to a different maximum point.
For the resulting firing rate f the equation applies
![]()
The following equations apply to the vectors r1 to r4 :
![]()
![]()
![]()
![]()
Substitution yields the initial equation
![]()
![]()
We exclude common factors.
![]()
Now we insert the quantities f1 to f4 . We had already obtained equations for these:
![]()
![]()
![]()
![]()
We use and maintain this:
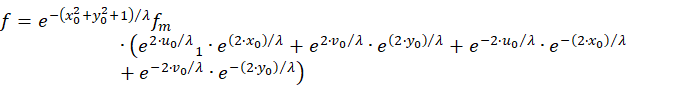
![]()
We apply the following addition theorem.
![]()
This gives us the functional equation
![]()
We set u + v = w, u - v = z, x + y = s and x - y = d and obtain because of
![]()
A new equation in the variables s and d instead of x and y:
![]()
We want to determine for which values of s and d the function value of f assumes a maximum. To do this, we form the partial derivatives according to s and d and set them equal to zero. We note that the factor fm contains neither the variables x or y and thus also not the variables s and d, i.e. it represents a constant in the sense of differential calculus.
![]()
![]()
We note that the factor
![]()
is because fm and λ are always positive and greater than zero and so is the exponential function, also the function cosh(x) is always greater than zero. Likewise
![]()
Thus, setting the derivatives to zero yields the two conditions:
![]()
![]()
Simple forming yields the following conditions
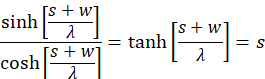
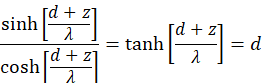
We apply the inverse function to tanh(x) and obtain:
![]()
![]()
![]()
![]()
Because of put u + v = w, u - v = z, x + y = s and x - y = d now also holds:
![]()
![]()
We can express u and v by t and ω.
![]()
![]()
![]()
![]()
About
![]()
Applies now:
![]()
![]()
For this angle ω, the neuronal excitation assumes a maximum.
We can express x and y by r and α.
![]()
![]()
![]()
![]()
Then we obtain a different representation for the phase particle ω.
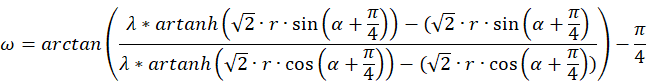
And last but not least, we can express the function Artanh(x) by the logarithm.
![]() for amount(x) < 1.
for amount(x) < 1.
Then applies:
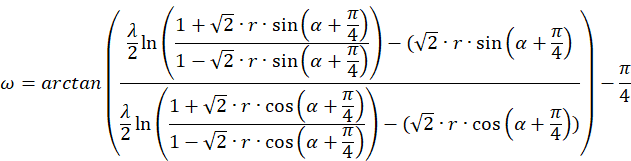
We now try to calculate the radius vector t.
About
![]()
also applies
![]()
![]()
![]()
If we replace x+y and x-y with the derived equations, we get the following result:
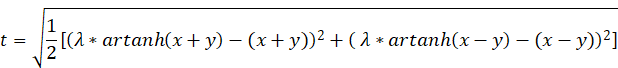
If the function f(x,y) is to assume a global maximum at the point x and y, the values u and v, respectively t and ω, must fulfil the above conditions.
Thus, there is a clear connection between the direction of gaze of the eye and the location of the excitation maximum in the cortical projection field of the signals of the tendon organs of the eye.
The formula for the phase angle ω looks very complicated, but every mathematician recognises that it is a periodic function of the angle α in the control plane of the eye muscles.
If the original radius vector t rotates through an angle ω in the field of view, the radius vector r also rotates through an angle α in the cortical field.
Thus, if a small black object in the field of view rotates around the centre of the field of view with gaze following it, the tendon organs of the eye muscles generate an output in the cortical projection field and the excitation maximum also rotates around the coordinate origin and is characterised by the radius vector r unn the angle α.
Using the programme Excel and the possibility it contains to create spatial representations of functions of two variables, we can graphically represent the radius vector t as a function of the extreme point P(x,y).
The function progression of t can be shown once viewed laterally from above and slightly rotated, as indicated here.
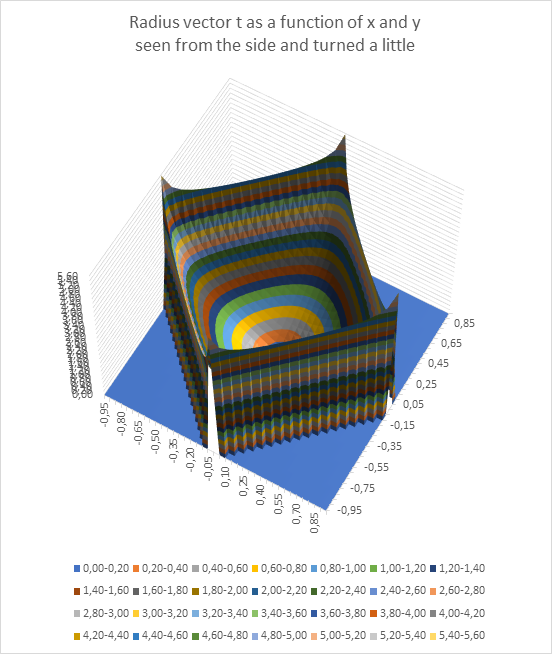
Here you can see that only values of t that lie in a rotated square lead to a maximum of the excitation at all. The blue area contains the impermissible values of x and y, in which no maximum can occur at all.
The function surface resembles a square bowl whose lowest point is exactly in the middle at x = 0 and y = 0 and which rises from the middle to the edge.
The phase angle ω can also be represented as a function of x and y. Each angle ω corresponds to a different colour (see legend).
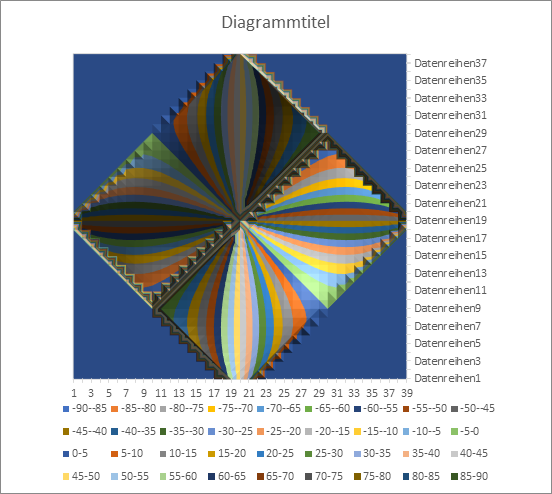
You can see a similar representation to the orientation columns in the visual cortex. No maximum can exist in the area coloured blue. In the remaining area, you can see that the angle ω takes on windmill-like values between -90 and + 90 degrees. If the radius vector t rotates around the centre point, the point P(x,y), where the maximum excitation in the x-y plane is assumed, also rotates. However, the non-linearities of the transmission characteristics of the neurons cause the uniform rotation of t to be transferred into a somewhat uneven rotation of the vector r. Only at multiples of the angle of 45 degrees do the directions of t and r coincide; in between, r runs slightly ahead or behind.
Now we need to discuss eye control in more detail. The visual input field in the tectum - of which there are independent input fields for different visual modalities - is used for eye muscle control. An object that lies exactly in the centre of the visual field generates equally strong signals in all four projection neurons in the tectum to the eye muscles or the neck muscles, so that all muscles contract equally. As a result, the eye does not move.
If the object is not central but eccentric, the eye muscles receive signals whose strength increases with increasing eccentricity. The contraction of the eye muscles causes the eye to move in such a way that the gaze is centred on the object. Since the object is now in the centre of gaze, the muscle signals are now identical. Since the muscles work like stepper motors, the bkick remains focused on the object.
However, it is very important for the creature to know exactly what position the eyes are now in. For this purpose, the muscle tension is measured with the help of the tendon organs, which fire more strongly the greater the muscle tension.
The four signals from the four eye muscles involved (two eye muscles serve to roll the eyes and are immaterial here) also reach the frontal cortex on its sensory side. There they contact the corner neurons of a "sensory" eye field that analyses the muscle contraction. These also form the corners of a larger quadrilateral that contains a myriad of output neurons. From the four input neurons, the excitation reaches the area of this sensory eye field and forms an excitation maximum by superimposition at one point, which also moves back and forth with the observed object when the eyes move, since the movement of the maximum is coupled to the movement of the object.
But what does the brain do with this information?
What does the brain gain from the fact that an excitation maximum wanders around in the sensory field and "encodes" the location of the visual object, so to speak. Every location corresponds to a location in the visual field.
The benefit is that this senssory eye field activates the living being's memory. This field practically logs the currently present system states. Therefore, we will call this type of cortical field, which receives input from tendon organs and converts it into maximum coding as divergence modules, sensorimotor protocol fields.
The active output neuron of the sensorimotor protocol field in the frontal cortex activates a neuron in the nucleus ruber, which activates a neuron in the nucleus olivaris. The axon of the latter activates a Purkinje cell in the cerebellum. If the object moves in the visual field, another cortical neuron and ultimately another Purkinje cell is strongly activated by another climbing fibre.
At the same time, a signal projection occurs in the eye from the area of sharpest vision - this represents a section of the visual field - via the bridge nuclei into the moss fibres and ultimately into the parallel fibres of the cerebellum. The excitations caused by the visual signals of the object propagate along the parallel fibres and reach the relevant, strongly activated Purkinje cell. Through the process of long-term depression (LTD) and long-term potentiation (LTP), the synaptic coupling strength of the active parallel fibres is abruptly changed so that this Purkinje cell stores the current image of the object and is later able to recognise it.
The output of the divergence fields that receive the muscle tension signals is therefore used to store (log) the signals that are currently present. In the optical case of storage, these are visual images. As soon as the eye directs its gaze to a certain point in the field of vision, an associated Purkinje cell is activated and stores the applied signals and thus the current image.
Now it becomes understandable why we observe a strong signal divergence in the divergence fields or divergence modules. The more output neurons there are, the more accurate the data stored in the Purkinje cells.
Since all joints use tendon organs to measure muscle tension and project to the cortex, there is such a sensorimotor protocol field for each joint. And each one stores, at each moment, the creature's ACT signals, which are able to reach the parallel fibres of the associated cerebellum cluster and its neighbouring clusters (cluster group).
These are at least the signals of all the other tendon organs, so that the living being constantly has an up-to-date model of the motor function of its body. This is how body consciousness could arise. In each of the many individual sensorimotor protocol fields, exactly one neuron is maximally excited and represents the joint angles of the joint; its output links all the current joint angles of the body together in the cerebellum. Thus, an active neuron cloud, diffusely distributed in the cerebellum, could be the representative of the motor body consciousness. The binding problem was already discussed in detail by Prof. Wolf Singer. It seems possible that sensorimotor protocol fields could be responsible for the binding of motor perceptions.
Monografie von Dr. rer. nat. Andreas Heinrich Malczan