Human brain theory
for the European Union’s Human Brain Project
ISBN 978-3-00-068559-0
14.7 The module for stereoscopic vision
Chapter Summary
This chapter introduces a neural module that constructs a topological map from the signals of the four rectus muscles of both eyes. This internal map forms a coordinate system in which one axis represents the average gaze direction and the other the distance to a visual object. The current viewing position generates a peak of neural activity in this map, which is stored in the cerebellum via long-term depression (LTD) as a visual memory trace. In this way, the brain maintains a dynamic internal representation of where visual objects are located and how far away they are.
Vertebrates have two eyes. This enables them to see spatially. Therefore, they must have a module in their brain that enables spatial vision. Perhaps several modules are necessary for this. This is because several subsystems are involved in the visual process. On the one hand, eye muscles control the movement of the eyes. Furthermore, there must be a control for the pupils. And last but not least, the shape of the eye lens has to be changed depending on the distance of the targeted object so that this object is imaged on the retina with the best possible sharpness. But the pupils must also be controlled in order to adapt the eyes to the existing average brightness.
But the visual system has other functions. For example, it is necessary for predators to determine the distance to the object so that one can estimate whether it is a possible prey or can no longer be reached due to its great escape speed.
Equally important is determining the direction to the object. This becomes even more complicated with those birds of prey whose prey are also flying birds.
And ultimately, the visual signals form the basis for the sense of orientation. What algorithms might underlie the fact that a hamster finds its own burrow again after foraging in the field?
There must be subsystems in the vertebrate and human brain that can handle all these tasks. The author will show that the basis for all these abilities is laid by ordinary divergent modules, with their output being further processed in both the cerebellum, the basal ganglia and ultimately the cortex.
We will not be able to describe all visually effective modules. However, it must be pointed out that these descriptions are initially hypothetical. The proof that they exist in reality must be provided by others. Here, we merely point out which algorithms nature could use to be able to achieve such goals.
Starting from the early rope ladder nervous system, we divide the neuronal nuclei into sensory nuclei and motor nuclei, these are located on both halves of the creature's body. In addition, there are sensory and motor side-changing nuclei.
The visually effective nuclei, which predominantly serve to control the eye muscles, are located in the segmented cord ladder system or its successor with certainty in the tectal floor.
In the chapter "Movement control by focal modules" it has already been shown how the sensory tectum opticum works as a focal module and generates the signals for the four straight eye muscles with which the gaze can be controlled both laterally and in height. Likewise, these tectal output signals are suitable for controlling the neck muscles, partly also for controlling the trunk muscles (trunk rotation for visual object tracking) or even for controlling arm movements, for example to grasp an object.
But how can we explain spatial vision and the ability to form a spatial image of the environment, in which the recognised objects are stored in a kind of map in terms of their position and distance?
Such a map is a current image of the environment. As soon as the living being changes its location, the previous map is invalid and a new one is created.
However, there is also a neuronal map that is preserved when the creature changes location, because the creature's location in the map changes as soon as it moves.
And finally, there is also a neuronal map that maps the environment, but in such a way that the map turns as soon as the living being turns. Just like a map in a GPS-controlled navigation device does. We will probably not get that far in this monograph, as the neuronal algorithms required for this have not yet been fully understood by the author.
We will design here - probably for the first time - a visual module for spatial vision and show where it is located in the brain. Here, the brain architecture should be taken into account as realistically as possible. The major shortcoming of many brain theories is that they neglect the real structure of the brain and the nervous system and approach the problem purely theoretically.
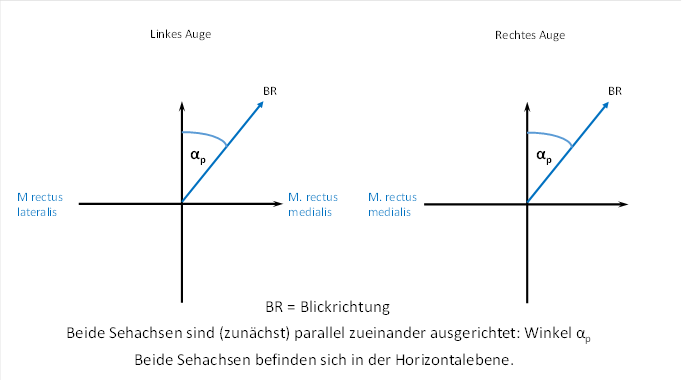
We first discuss the case where the visual axis of both eyes is in a horizontal plane while the head is in the normal position. We therefore look horizontally forwards or slightly to the side without raising or lowering our gaze.
First, we look straight ahead into the distance, virtually into infinity. Both visual axes of both eyes are aligned parallel to each other.
Now we turn our gaze slightly to the right and also look into the distance, so that this time, too, both visual axes are aligned parallel to each other.
Here the angle αp occurs. It is supposed to be the angle that the visual axis makes with the line of sight when we look straight ahead. This can be seen in the following figure. We call the angle αp , because it refers to distance vision, where the visual axes of both eyes are aligned parallel to each other.
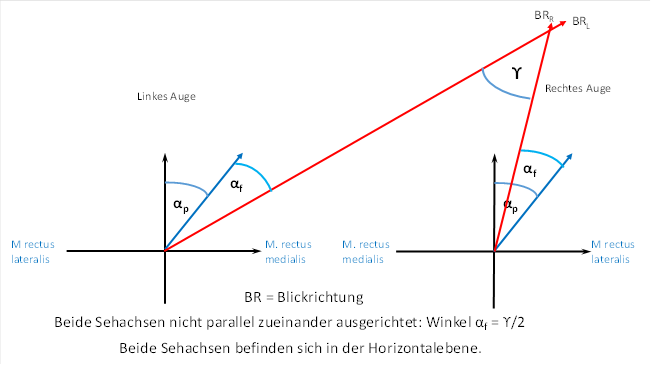
Now we decide to fixate with our eyes on an object that is on the right side and only a few metres away from us. Here, the two visual axes are no longer parallel to each other. For the left eye, we have to increase the angle αp by an additional angle αf . We call the additional angle αf , because it is necessary when focusing on a nearby object. For the right eye, we need to decrease the angle αp by one the angle αf . This shows that the angle αf is exactly half the angle ϒ. The angle ϒ is the intersection angle of both visual axes.
This is shown in the figure below.
We denote the deflection angle from the straight-ahead position for the left eye by αL and for the right eye by αR . This is shown in the following figure.
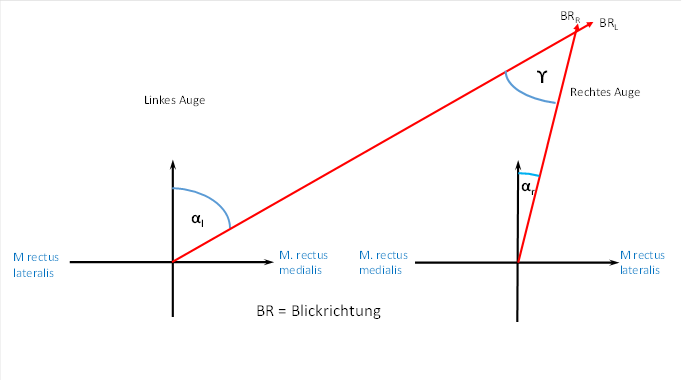
A comparison of the two representations yields the formulas we are looking for for the direction of gaze.
![]()
![]()
![]()
We now need a theoretical approach for the firing rate of the eye muscles involved. We assume an average firing rate fm . This should be present when the eyes look exactly straight ahead without raising or lowering the gaze.
If we look to the right in the direction of gaze given by the angle α, the eye muscle that directs the gaze to the right must contract more strongly. For the left eye, this is the musculus rectus medialis, for the right eye, on the other hand, the musculus rectus lateralis. The respective motor counterpart, on the other hand, must reduce its contraction somewhat.
Concrete formulas for the contraction of the eye muscles are missing in the previous theories. We assume that the firing rate grows exponentially with the angle of deflection, accommodating the longitudinal constant λ of the neurons in the formulae.
We use the index L for the lateral rectus muscle and the index M for the medial rectus muscle.
Then let the following approach be used for the firing rates f:
|
Musculus rectus |
Rate of fire |
|
|
left |
medialis |
|
|
left |
lateralis |
|
|
right |
lateralis |
|
|
right |
medialis |
|
The muscle that is more tensed has the plus sign in the exponent. The muscle that is less tense can be recognised by the minus sign in the exponent.
We see that in both eyes the firing rate of the rectus lateralis muscle is inverse to the firing rate of the rectus medialis muscle. Both are to be classified as motor antagonists to each other.
From these four fire-rate signals, we want to construct a module that enables spatial vision and that actually occurs in a real structure of the brain.
But before that, we need to remember the two other types of eye muscles, as their signals also need to be considered. They are the superior rectus muscle and the inferior rectus muscle. The first raises the gaze, the second lowers it. We assume that the gaze is raised by the angle β. This angle is identical for the left and right eye.
Then let the following approach pass the connection between the angle and the rate of fire:
|
Musculus rectus |
Eye left |
Eye right |
Effect |
|
superior |
|
|
Lift gaze |
|
inferior |
|
|
Lower gaze |
This gives us 8 eye muscle signals, four from the right eye and four from the left eye. Now we only have to clarify in which way these signals could be arranged in a module or a neuronal nucleus.
Since signals from both sides of the body are involved, there is only one type of core that processes both signal versions. This core must be a side-switching core.
Even in the primitive rope ladder nervous system, there was a motor and a sensory side-changing nucleus on each floor and on each side of the body. The corresponding signals from one side of the body travelled through them to the opposite side. This enabled the exchange of signals between both sides of the body.
The signals from the ipsilateral side of the body moved to the side change nucleus to get to the opposite side. Similarly, the signals from the contralateral side moved to the side change nucleus to get to the ipsilateral side. Thus, there is a meeting of these signals here, so that neuronal structures could access both sides of the body.
There are both sensory and motor side change nuclei. Here we examine the motor side change nuclei for the signals of the tendon organs of the straight eye muscles of both eyes.
The left eye signals travel to the left lateral nucleus, the right eye signals to the right lateral nucleus. Both are located on the motor side of the old cord ladder nervous system or neural tube in the floor containing the tectum opticum, which evaluates the eye muscle signals.
If one imagines this spatially in the mind as a model, one notices the close proximity of both nuclei.
Nature has certainly arranged it so that the two different motor side-changing nuclei of the two sides of the body are so closely adjacent that they could eventually merge in the course of evolution. Then there is only one common motor side-changing nucleus in each segment of the segmented nervous system, which receives input from both the left and right sides of the body at the same time.
We can imagine this common motor side-switching nucleus - which is certainly located in the tectal floor - as a cube. The four signals of the left eye end at the left face of the cube in the four corners. The signals of the right eye end correspondingly in the four right corners of the cube.
For further analysis, we turn the cube in such a way that the signals for the horizontal eye movement also lie in the horizontal plane. This facilitates the later observations.
Further, we turn the cube so that the two corners that receive the signals from the muscles that lift the gaze are at the top. The other two corners receive the signals from the muscles that lower the gaze and are at the bottom.
This gives the cube the position we need for our analysis.
In the centre of the cube field we place the origin of a spatial coordinate system. Let the x-axis run from left to right in the horizontal plane, the y-axis from front to back and the z-axis from bottom to top. Let the neuronal signals of the eight eye muscles with their firing rates f1 to f8 arrive in the eight corners of the cube.
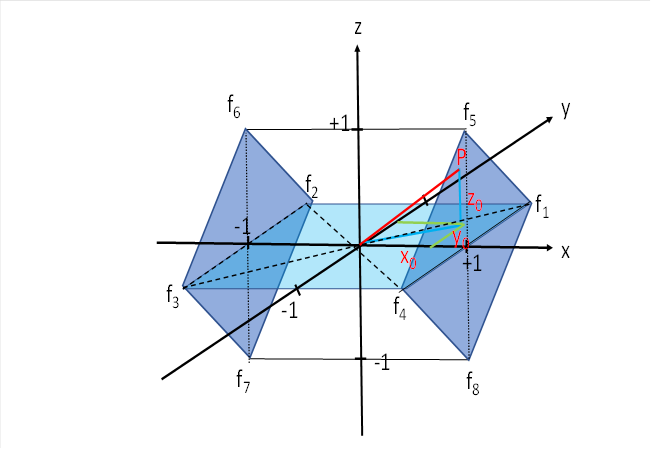
We will first examine only four of the eight signals, limiting ourselves to the eye muscles that cause eye movement to the right and to the left. The muscles that cause the gaze to be raised or lowered will be ignored for the time being; we will only include them in our consideration later. So for now we will limit ourselves to the square, which lies in the x-y plane. It is light blue in the above illustration.
The input of the selected four eye muscle signals for the lateral eye movements should arrive at the four points P1 (1;1), P2 (-1;1), P3 (-1;-1) and P4 (1;-1). This is shown in the figure below. It shows the x-y plane we are referring to.
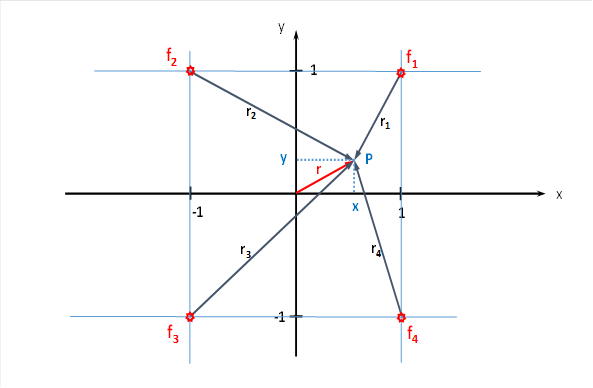
For the total excitation at the point P(x,y) the firing rate then applies
![]()
These four neuronal excitations spread out in the x-y plane in all directions, and the transmission of excitation is realised by the countless interneurons in this structure. Thus, a divergence module is present here. Here - as is usual in a divergence module - an exponential signal attenuation occurs, which increases quadratically with the distance. With the longitudinal constant λ of the interneurons involved, we can calculate the excitation arriving at the point P(x,y).
If a point P is at a distance r from an input neuron with a firing rate f0 , the relationship between the input excitation f0 and the excitation at point P is described by the following formula:
![]()
Here λ is the longitudinal constant of the interneurons.
In the exponent, the distance occurs quadratically because the excitation no longer spreads on a single axon but via strongly branched axon clusters. As a result, the excitation decreases faster with distance than on a straight, unbranched axon.
This excitation arriving at the cube corners is picked up by a neuron at point P (or in its immediate vicinity) and represents the output of the divergence modulus at point P(x,y).
Since the individual partial summands of the equation
![]()
represent strictly concave functions in the variables x and y in the square under consideration (keyword Hessian matrix), a global excitation maximum exists in this area, which neurologists could possibly detect by measurement. If the position of the eye angle changes, this excitation maximum would move around. We are interested in which eye corners can be assigned to an excitation maximum. Is it possible to determine the angles αp and αf of the eyes from the coordinates x and y of the excitation maximum?
Here we have to apply differential calculus by calculating the partial derivatives of f(x,y) with respect to x and y and setting them equal to zero.
We first calculate the squares of the radius vectors.
![]()
![]()
![]()
![]()
We substitute the squares of the radius vectors into the initial equation, but subtract the common factor before the sum.
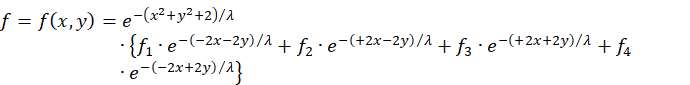
![]()
Now we set the firing rates f1 to f .4
We only have to determine which of the four signals should arrive at which point. We decide to place the two signals of the eye muscles that cause the gaze to move to the right at the top. The signals that cause the eye to move to the left are placed at the bottom.
In this case, the signals of the left eye are generally to be arranged on the left, those of the right eye on the right.
Then applies:
![]() right eye, muscilus rectus lateralis,
view to the right
right eye, muscilus rectus lateralis,
view to the right
![]() left eye, musculus rectus medialis,
view to the right
left eye, musculus rectus medialis,
view to the right
![]() right eye, musculus rectus medialis,
view to the left
right eye, musculus rectus medialis,
view to the left
![]() left eye, musculus rectus lateralis,
view to the left
left eye, musculus rectus lateralis,
view to the left
The reason for this special arrangement is based on the assumption that there is a body side marker. Therefore, the input neurons for an eye movement to the right are located on the right side of the square. Similarly, the input neurons for an eye movement to the left are located on the left side. The body side marker is transmitted from the muscles concerned via the tendon organs to the target neurons in this tectal structure.
The insertion of the fire rates delivers:
![]()
![]()
Summarising provides an intermediate result with hyperbolic functions according to
![]() resp.
resp. ![]()
This results in:
![]()
We apply the following addition theorem for hyperbolic functions:
![]()
The application of this addition theorem yields:
![]()
![]()
About
![]()
![]()
and
![]()
![]()
results in the final formula:
![]()
This formula describes the total excitation at the point P(x,y) of the tectal lateral motor nucleus when the excitation is restricted to the x-y plane.
Due to the strict concavity of this function in the considered square with side length 2, a global maximum exists there.
By zeroing the partial derivatives to x and to y, one can derive conditions for the global maximum.
We calculate the partial derivative with respect to x.
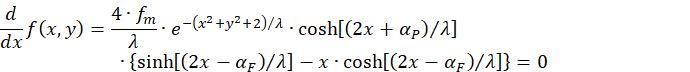
About
![]()
the following condition must be fulfilled for the partial derivative with respect to x to be equal to zero:
![]()
This is equivalent to
![]()
We reshape:
![]()
![]()
Thus we have obtained the first condition for the existence of a maximum of the excitation function f = f(x,y) at the point p(x,y), it reads:
![]()
A second condition arises if we set the partial derivative of the function with respect to y equal to zero:
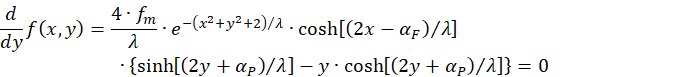
Again:
![]()
Thus, the only remaining condition for a maximum is the equation
![]()
transformation yields the second condition for the existence of a maximum:
![]()
Final result:
The excitation function
![]()
of the tectal motor side change nucleus in the x-y plane assumes a global maximum at the point P(x,y) if and only if the involved eye angles αF and αP satisfy the following conditions:
![]()
![]() .
.
Notice:
In the x-y plane of the tectal
side change kernel, the x-value of the global maximum encodes the angle of
focus ![]() and the y-value the angle of view
and the y-value the angle of view ![]() when looking into infinity.
when looking into infinity.
Since the focal angle is directly coupled to the distance of the targeted visual object, the x-coordinate of the global excitation maximum encodes the object distance of the targeted object.
But what benefit does the living being derive from the fact that in the tectal x-y plane of the lateral exchange nucleus, the object distance in the x-direction and the (average) angle of view (when looking into infinity) in the y-direction are encoded in maximum code?
Such an expenditure of resources must have brought advantages, otherwise such a system would not have emerged. The development was quite simple. The number of interneurons had to increase, and at the same time additional output neurons had to receive the transmitted excitation. And the additional interneurons and output neurons had to be distributed as evenly as possible in the module. The rest was done by the neuronal signal damping.
We repeat:
Even if we restrict ourselves to the consideration of the x-y plane, a divergence module is present here. In addition to the four input neurons at the corners of the square in the x-y plane, there are innumerable projection neurons distributed in the plane (and in space) within this plane. In between, there are countless interneurons in this square. They transmit the excitation from the four input neurons to the hundreds or even thousands of projection neurons. Obviously, there is also a transmission of excitation from interneuron to interneuron, which are quite small compared to the total extension.
Here, a distance-dependent damping occurs which increases quadratically with the distance between the exciter and the excited. This transfer function is therefore strictly concave within the square and in its vicinity, i.e. always curved upwards as a function surface. Therefore, there is exactly one excitation maximum in the surface. This is extremely important!
Before the output leaves this structure, it must (most likely) pass through a layer of inhibitory interneurons. These provide lateral inhibition for contrast enhancement. This is common in neuronal nuclei.
The output neuron that is located at the site of maximum excitation fires first and excites the inhibitory interneurons. These inhibit all the other output neurons of the structure before they themselves reach the threshold for triggering an action potential. Thus, only one output neuron fires in the module, the others remain silent.
Where does the output of all the projection neurons in the tectal x-y area go?
Since it is a motor side change nucleus, it is located on the motor side of the old cord ladder nervous system or neural tube. There, signals descend on the axons of class 5 neurons. Motor signals reach the motor neurons of the muscles to be controlled via the nucleus ruber.
But the output of the tectal, motor side-changing nucleus has (with one exception!) no motor targets. Once it reaches the nucleus ruber, it only has the path to the nucleus olivaris, which sends these signals via the climbing fibres to the cerebellum. (One exception is presented at the end of the chapter).
Thus, in the cerebellum, or more precisely in the pontocerebellum, there is an analogously organised square of Purkinje cells (or more precisely Purkinje groups) in which the output of the tectal motor lateral alternating nucleus terminates. We call this area the cerebellar image of the tectal lateral nucleus.
The target site is certainly in the pontocerebellum, because the spinocerebellum only processes motor signals, which are not present in our case.
This is where the memory work of the pontocerebellum begins. Exactly one projection neuron in the tectal motor lateral nucleus is always maximally excited (because there is only one global maximum). The maximum represents a visual object that is fixated with both eyes. Above all, however, this maximum represents the object distance and the middle eye angle to the targeted object.
While the site of sharpest vision creates an image of the object on the retina when focusing, the visual signals of this image section reach the primary visual cortex via the visual thalamus.
From there, there is a projection via the bridge nuclei into the parallel fibre system of the pontocerebellum. We assume that the visual signals of the retina in their entirety reach exactly the cerebellar image of the tectal lateral exchange nucleus via the parallel fibres. For the sake of simplicity, we assume that only the site of sharpest vision has a connection to the cerebellum. If one directs one's gaze at any visual object in the horizontal plane, the eyes must catch it with the gaze, for which they must look to the left or to the right or even straight ahead. Depending on the distance of the object, the angles of the left and right eyes are usually different.
Now the analysis of the space in the plane begins (we have not yet included the influence of raising and lowering the gaze here, so we are only looking in the horizontal plane).
The analysis is carried out through the interaction of two algorithms.
-In the tectal motor lateral exchange nucleus, a projection neuron is
maximally excited, whose x-coordinate represents the focal angle ![]() and
whose y-value indicates the angle of gaze
and
whose y-value indicates the angle of gaze ![]() when
looking into infinity. This strong excitation reaches exactly one Purkinje
group in the area of the cerebellar image of the tectal
lateral exchange nucleus via the nucleus ruber and the nucleus olivaris.
This purkinje group is strongly excited and initiates the imprinting process in
which, through LTP and LTD, the parallel fibres that are currently active
undergo and permanently maintain a strong change in their synaptic coupling strength.
when
looking into infinity. This strong excitation reaches exactly one Purkinje
group in the area of the cerebellar image of the tectal
lateral exchange nucleus via the nucleus ruber and the nucleus olivaris.
This purkinje group is strongly excited and initiates the imprinting process in
which, through LTP and LTD, the parallel fibres that are currently active
undergo and permanently maintain a strong change in their synaptic coupling strength.
-From the area of sharpest vision, all visual signals reach the cortex via the visual thalamus and from there, via the bridge nuclei, precisely the parallel fibres of all Purkinje groups located in the area of the cerebellar image of the tectal lateral exchange nucleus . All the Purkinje groups there receive the current signal image of the work of the sharpest vision via the parallel fibres.
-But only exactly one of these Purkinje cells is excited via the climbing fibres from the tectal motor lateral exchange nucleus. Only this Purkinje group stores the current signal image from the area of sharpest vision.
-The other Purkinje groups do not receive any climbing fibre excitation from the tectal motor side-changing nucleus and therefore cannot store the current signal image. This is because in the tectal motor side-changing nucleus the output is subject to neuronal competition. Inhibitory interneurons ensure that only the output at the point of maximum excitation prevails. It destroys the existing, significantly lower excitation of the other output neurons. But if the gaze in the past was directed at an object whose angular values belong to the silent Purkinje group, then the image of the object observed at that time is mapped in this Purkinje group.
-However, since the eyes (unconsciously?) "scan" the entire environment during vision, this system creates a complete image of the environment, which, however, consists of a number of small partial images that our brain perceives as a complete image. If the gaze wanders back and forth, this complete image remains unmoved, only the focus of attention wanders with a change in the direction of gaze to the point of the complete image that is represented by the corresponding partial image in the cerebellum. This gives us the impression that the world is standing still and only our gaze wanders back and forth (as long as the head does not move).
Thus, for every direction of gaze actually analysed, there is a visual image in the cerebellar image of the tectal lateral exchange nucleus. This is the reason why we perceive the environment in such detail and can remember what we saw where and in which direction, even if we close our eyes for a short time. In our subconscious, the pontocerebellum, or more precisely in the cerebellar image of the tectal lateral nucleus, each individual Purkinje group has mapped a partial section of the visual environment once we have looked in the corresponding direction and distance. And we do this automatically. We search the environment for objects and store their visual image in the Purkinje cells of the pontocerebellum within seconds.
And the inverse pontocerebellum can reactivate the stored object images in consciousness by sending them back to the visual thalamus as soon as the associated Purkinje group is activated. This process is somewhat more complicated and will not be discussed here.
In summary, the tectal motor lateral alternating nucleus provides the basis for our spatial perception.
Addition dated 20.10.2023:
Now recall that there is also signal divergence in the nucleus olivaris. This means that the currently active neuron in the tectal lateral alternating nucleus projects into several neurons of the nucleus olivaris. Let us first assume, in a simplified way, that these are connected in series, so that there is a sequential divergence here. Then several different images could be stored for a given line of sight. It would therefore be technically possible to store different object images and to recognise them later. In the case of a hamster looking for its way home, these could be distinctive objects along the path that mark the way to the hamster's den. This leads to the conclusion that this neuronal form of organisation must already be present in simpler vertebrates.
Until now, however, a restriction was made: the eyes were only allowed to look out in the horizontal plane. We are now dropping this restriction. Now we investigate the functioning of the tectal motor side-changing nucleus under the condition that the gaze of the eyes can also be directed upwards or downwards. Then a spatial model of the environment is created in the tectal motor side-change nucleus, in which each recognised visual object can be assigned its spatial coordinates. This module thus creates the basis of our spatial perception.
We look again at the illustration with which we started this chapter.
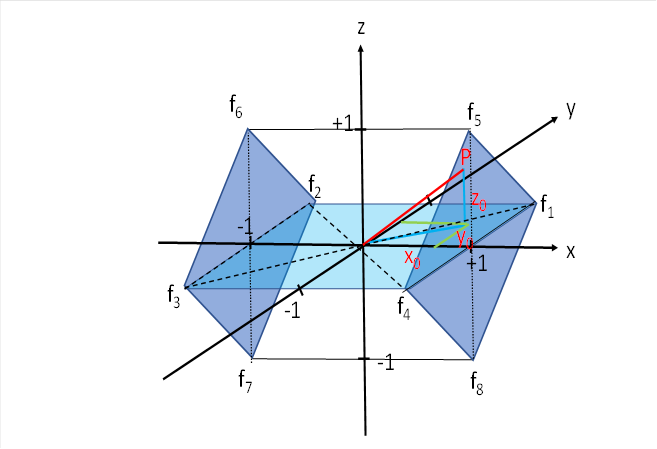
So far we had only considered the square, which was in the x-y plane. For the sum of the four excitations, the following had already been derived in the chapter "The motor divergence module with lateral signal propagation":
![]()
![]()
When we leave the plane and go into space, we have to consider the variable z in addition to the variables x and y.
Since a spatial radius vector t must be supplemented from the radius vector r of the plane by the additional coordinate z, the new summation formula for the excitations in the plane is as follows:
![]()
The following applies here:
![]()
![]()
![]()
If you insert this into the above summation formula, you can pull the common factor in front of the sum:
![]()
About
![]()
also applies:
![]()
About
![]()
we obtain for the spatial excitation function in the tectal motor side alternating nucleus the equation
![]()
Now it is particularly easy to see that αp depends only on the variable y and and αf only on the variable x when it comes to the global maximum.
Since this is the portion of the excitation caused by the eye movements in the horizontal, we rename it and henceforth call it fH . Then holds:
The complete calculation of the partial derivatives of fH (x,y,z) results in the following three equations, which must be fulfilled as conditions for the existence of a global maximum at the point P(x,y,z):
-![]()
-![]()
-![]()
We see that the first two conditions are identical to those we had obtained for the planar tectal motor side exchange nucleus.
Four excitation components are now missing, which are delivered by the eye muscles that are responsible for raising and lowering the gaze. Their signals arrive in the upper and lower corners of the two darker blue squares.
We look at the two squares in the figure above, which lie parallel to the y-z plane (dark blue). The eye signals responsible for raising and lowering the gaze arrive in its four corners. At the top are those for raising the gaze, at the bottom those for lowering it. The signals of the left eye are on the left and those of the right eye are on the right.
We look again at the figure for the tectal motor lateral exchange nucleus.
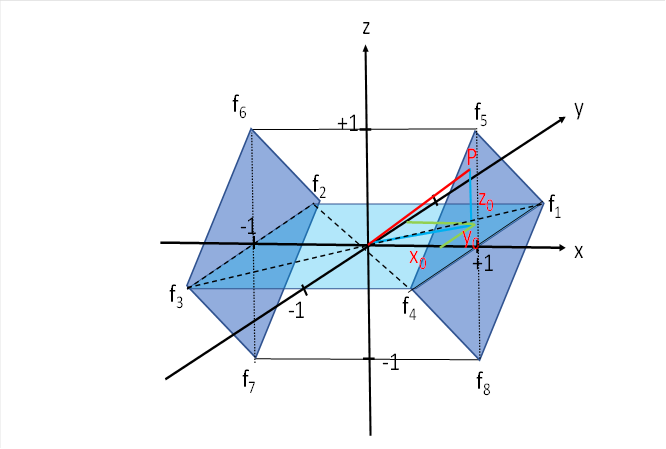
We now have to calculate the excitation component fV for the firing rates f5 to f8 , which enables the vertical eye movements. We denote by t5 to t8 the radius vectors representing the distances of the point P to the four respective corners of the cube where these four firing rates arrive.
In principle
![]()
The following applies to the fire rates concerned (see beginning of chapter):
|
Musculus rectus |
Eye left |
Eye right |
|
superior |
|
|
|
inferior |
|
|
Since the firing rates for the left and right eye are identical, we can already summarise here.
![]()
We first calculate the length of the radius vectors, where we need the squares of them right away.
To do this, we must assign points to the four corners and determine their coordinates in order to calculate the length of the radius vectors.
At the point P5 the firing rate f5 arrives, at P6 the firing rate f6 , to the point P7 belongs f7 and to P8 belongs f8 .
Then these points have the following coordinates:
|
Item |
x-coordinate |
y-coordinate |
z-coordinate |
|
P5 |
+1 |
0 |
+1 |
|
P6 |
-1 |
0 |
+1 |
|
P7 |
-1 |
0 |
-1 |
|
P8 |
+1 |
0 |
-1 |
The distance t of a point with the coodinates P(x,y,z) to any point with the coordinates P(a,b,c) is given by
![]()
The following squares of the radius vectors therefore result for the four
points:
![]()
![]()
![]()
We first calculate
![]()
![]()
![]()
![]()
We also charge
![]()
![]()
![]()
![]()
Substituting into the equation for fV yields
![]()
![]()
![]()
We first calculate the conditions for the presence of a maximum for this vertically effective excitation component in order to be clear about what influence it has.
To do this, we calculate the partial derivatives with respect to x, y and z and set them equal to zero.
![]()
This equation is only fulfilled for
![]()
It follows immediately
![]()
Thus the extreme value condition applies to the variable x:
![]() (derivative of fV according to x)
(derivative of fV according to x)
We calculate the partial derivative with respect to y and set it equal to zero to obtain another condition for the presence of an extreme value.
![]()
The following extreme value condition necessarily follows from this:
![]() (derivative of fV according to y)
(derivative of fV according to y)
Finally, we calculate the partial derivative with respect to z and set it equal to zero to obtain the third condition for the extreme value.
![]()
This equation can only become zero if the following holds:
![]()
![]()
![]()
![]()
![]()
Thus, for the existence of an extreme value, the third condition must also be fulfilled:
![]() (Derivation of fV according to z)
(Derivation of fV according to z)
We summarise:
The function fV (x,y,z) has a global maximum at the point P(x,y,z) if and only if the following three conditions are fulfilled (simultaneously)
- x = 0
- y = 0
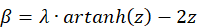
The extreme value is therefore exactly on the z-axis at the height given by the third equation. As z increases, the angle β becomes larger; as z decreases, this angle becomes smaller.
Or vice versa: the larger z is, the larger the angle β must become so that the extreme value lies at point z again.
However, we recall that these equations only apply to the excitation portion of the eye muscles, which include for the vertical eye movements (up and down).
Normally, the tendon organs of the eye muscles will contribute both to the horizontal excitation fH and to the vertical excitation fV , since in most cases the gaze is not directed rigidly straight ahead, but rather somewhat sideways as well as upwards or downwards.
In these cases, both excitation components overlap and add up. This shifts the maximum. Because of the concavity of the partial functions and the superposition result, the new maximum lies on a connecting line of the original two maxima that each of the partial functions had. Unfortunately, this connecting line is not a straight line, but curved. Therefore, the position of the new extreme value is very difficult to calculate.
We remember: The horizontal part of the excitation was
![]() .
.
The associated conditions for a maximum at the point P(x,y,z) were
-![]()
-![]()
-![]() .
.
The vertical portion of the excitation was
![]() .
.
The associated conditions for a maximum at the point P(x,y,z) were
- x=0
- y=0
Each of the two partial excitations already has a gradient field, which is not spherically symmetrical, but is somewhat distorted by the parameters of the eye angles.
While the horizontal component represents an excitation field that has a global maximum at the point PH (x,y,0), the excitation maximum of the vertical component is located exactly on the z-axis at the height z, i.e. at the point PV (0,0,z).
By superimposing both parts, the maximum in point PH (x,y,0) is pulled in the direction of point PV (0,0,z), but not in a straight line, but along the current gradients. However, these are (mostly) curved. Along the straight-line connection of both points, a maximum also exists because of the strict concavity of both parts, but it is only a local one along this straight line. The absolute maximum lies somewhere near this line, but its correct determination is extremely difficult. Determining the three eye angles αF , αP and β by setting the partial derivatives to zero seems impossible to the author after many attempts. May others dare to try.
As a result, however, it remains to be said:
In the spatial tectal motor side-exchange nucleus, the maximum excitation of one of the many output neurons represents exactly one spatial point that is targeted by the eyes. The maximum excitation arises in the module on the one hand from the input of the involved eight straight eye muscles of the two eyes, and on the other hand from the spatial propagation and superposition of these excitations in the three-dimensional space of the module. The signal attenuation during propagation increases quadratically with distance. This results in a strictly concave superposition function with a unique global maximum in the module.
The mapping of the surrounding space in the tectal motor nucleus is transferred to the cerebellum. Here, however, the spatial mapping must be transferred into a mapping in the surface, because the Purkinje cells form a surface. The cube (see illustration) must therefore be cut into slices, which are then arranged one after the other. Each slice represents a height of space, we will call it a height slice. As the area becomes narrower towards the top and bottom (the cube is rotated in space), the height slices also become narrower. Thus, the represented area in the cerebellum has the shape of an elongated rhombus that is much narrower than its total length.
This decomposition into slices is also the reason why the neuronal representation of a moving object in space in the cerebellum, but also in the subsequent hippocampus - suddenly shows jumps. If the object moves in such a way that there is a transition from one slice to another, the maximum suddenly jumps from one slice to the other. This is where a discontinuity occurs.
Each of these areas is sent headward via an ascending projection and reaches the first floor of the early cord ladder system, where it ultimately lands in the hippocampus. There one also finds the successively arranged elevation slices, of which the one for the horizontal plane has the greatest width.
If a visual object now moves back and forth in the plane, for example a swimming duck on a water surface, the image of this duck is also moved back and forth in the associated height slice. Along the duck's path, exactly one hippocampal neuron will always be maximally excited, because the associated neuron in the tectal motor lateral switch nucleus is maximally excited and the excitation reaches the hippocampus via the cerebellum. This contains an image of the surrounding visual space, and the neuron that is maximally excited by the signals from the tendon organs of the muscles in the tectal divergent module represents the targeted visual object - in our case the duck. If the image of the objectd leaves the current height slice due to a change in height, the excitation maximum jumps to the next current height slice - thus showing discontinuous behaviour.
This image of the tectal motor lateral nucleus should not be taken as a navigation device, because it is always a very quickly transient image of the momentary state of the eye muscles. It is therefore more like a map of the eye directions.
Nevertheless, the phenomenon will already occur here that the excitation always spreads along hexagons when the object - for example the duck - is moved. To be more precise, however, they are triangles, which in turn form hexagons.
Why is that?
The many thousands of axons that carry the signals from the cerebellum to the hippocampus can be interpreted as straight pipes of constant diameter. And when there is a lack of space, these arrange themselves in such a way that the tubes lie staggered on top of each other, creating a kind of honeycomb shape in the cross-section made up of lots of small triangles. Thus, the observed honeycomb shape in the hippocampus in the area of the representation of the eye directions has its cause in the space-saving arrangement of the axons, which later retain this honeycomb-like shape when fanning out in the area. Therefore, one does not have to look for neuronal networks that lead to the honeycomb shape of the active signals through "learning processes". There are often simple explanations in the nervous system.
The discovery of mirror neurons is another such case. When one duck looks at another swimming around on the water, the path of the moving duck can be seen in the hippocampus of the observing duck through maximally excited neurons that trace the path of the moving duck. No mirror neurons are present here. When neurons fire, there is always an explainable algorithm for why they fire. They are not able to "understand" the actions of the other person. In general, we should get out of the habit of ascribing intelligence to neurons and using formulations such as: The neuron recognises ..."
Neuronal circuitry and receptor input force certain neurons to fire maximally under certain conditions. Intelligent is then not the firing neuron, but the living being in whose nervous system all this takes place. Intelligence is a system property, it cannot be assigned to individual neurons.
At the end of this chapter, we answer the question of which signals change the shape of the lens of the eye so that it focuses on the targeted object on the retina.
The muscle whose contraction increases the curvature of the eye's lens and thus its refractive index is the ciliary muscle, which surrounds the lens in a ring and is attached to flexible zonula fibres in the eye. It consists of three muscles: the muscle of Brücker, the muscle of Iwanoff and the muscle of Müller. When Bridge's muscle contracts, the refractive power of the lens is increased, thus enabling near accumulation. Müller's muscle also causes an increase in the refractive power. The motor counterpart of these two muscles is the Iwanoff muscle. When it contracts, the refractive power of the lens becomes smaller, thus setting a visual acuity for distance vision.
We need two muscle signals to control accommodation: one supplies contraction signals to the Brücker and Müller muscles , the other signal is the inverse of this and is intended for the motor counterpart, the Iwanoff muscle.
This pair of signals is generated incidentally in the tectal motor lateral exchange nucleus.
We look again at the corresponding illustration.
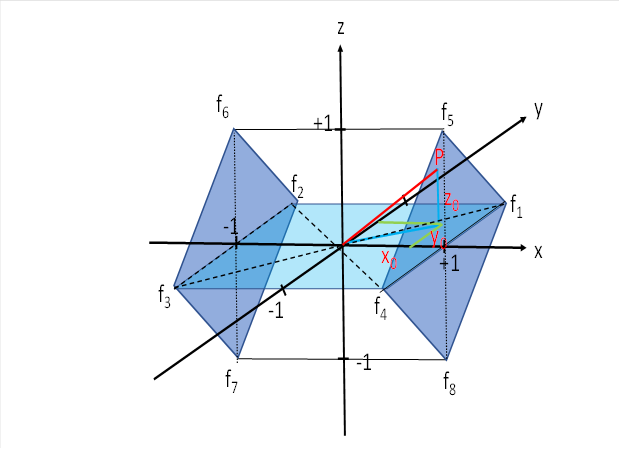
We need two signals for each eye. The following signals are required for this:
![]() right eye, muscilus rectus lateralis,
view to the right
right eye, muscilus rectus lateralis,
view to the right
![]() left eye, musculus rectus medialis,
view to the right
left eye, musculus rectus medialis,
view to the right
![]() right eye, musculus rectus medialis,
view to the left
right eye, musculus rectus medialis,
view to the left
![]() left eye, musculus rectus lateralis,
view to the left
left eye, musculus rectus lateralis,
view to the left
We recall the following contexts:
![]()
![]()
Then the following equations also apply:
![]()
![]()
![]()
![]()
We only consider the signals in the horizontal plane (light blue). First we form the quotient of f1 and f2 .
![]()
This firing rate quotient only contains the focussing angle αF . This angle is the difference between the eye direction of the left and right eye, it is necessary for focusing. Therefore, the two quantities f1 and f2 are suitable for controlling the muscles for the refractive power adjustment of the eye. The firing rate f1 is suitable for the Brücker and Müller muscles, and the firing rate f2 is responsible for controlling the counterpart, the Iwanoff muscle.
Because f1 increases with increasing angle αF , while f2 increases with decreasing angle αF . And since only the quotient of the two is important for muscle tension (extreme values remain unchanged when the output functions are multiplied by constants), the angle αP is not considered.
This makes the firing rate pair (f1 ;f2 ) suitable for controlling the ciliary muscle with its three components. And this signal can be used for both the left and the right eye, thus controlling the distance-dependent accommodation of the eye lens.
Thus, the tectal motor lateral exchange nucleus also performs the task of focusing the lens of the eye.
Monografie von Dr. rer. nat. Andreas Heinrich Malczan