Human brain theory
for the European Union’s Human Brain Project
ISBN 978-3-00-068559-0
17. The double layer theory according to Andreas Malczan
The double layer theory according to Andreas Malczan
This was originally intended to be a chapter on the pyramid of consciousness in the brain. But then this topic took on a life of its own, there was a paradigm shift and a separate chapter was developed as a result.
Many brain researchers are driven by the question of what differences there are between the brains of different classes of vertebrates. A very popular question is what differences there are between the brains of reptiles, birds and mammals. Curiously, one important vertebrate class is missing from this list: fish. This is despite the fact that they existed long before the other vertebrate classes.
Nevertheless, this question is obviously very interesting.
This is exactly the question that has been bothering me for decades! It seems that I have gradually come closer to the answer. And only recently, on 4 October 2024, I noticed in the publication "Purkinje cell neurotransmission patterns cerebellar basket cells into zonal modules defined by distinct pinceau sizes" by Joy Zhou, Amanda M Brown, Elizabeth P Lackey, Marife Arancillo, Tao Lin and Roy V Sillitoe that there is now experimental confirmation of my double-layer theory of the vertebrate brain. But to do so, one must recognise and understand the neuronal circuitry of the cortex and cerebellum as I have already described it in my monographs.
I've actually been interested in circuits of all kinds from the very beginning. As a teenager, I inspected the broken washing machine that my mother was given by her brother when she moved from Poland to Germany in 1957 and was allowed to take with her. However, it arrived broken, as did the large radio. The duvets had also been slashed open. Apparently, some railway officials had something against the emigrants or the immigrants.
At some point, around the sixth grade, I decided to repair the washing machine and the radio. In the library in Bad Doberan, our new home town, there were countless books on electrical circuits. In the GDR, the thirst for knowledge was very much encouraged, especially among working-class children. The radio started playing again after I had replaced the stolen tube. By then I already knew how radio waves were generated, how an audio receiver worked and why superheterodyne receivers were better. I began to take an interest in electronics and later I wanted to become a radio amateur, attended the necessary courses and even got a call sign.
During my studies, I acquired extensive knowledge of the natural sciences, but also of electronics in particular. I came to the conclusion that, with a sufficient level of knowledge, you could recognise and understand how any electronic circuit works at if you only knew the circuit diagram. There was plenty of evidence to support this assumption. For example, it was possible to recognise the functioning of German radar equipment when such equipment fell into the hands of the British during the Second World War.
I also came to the conclusion that the functioning of the brain could be deciphered by focussing on the brain's circuitry. After all, the brain processes signals and neurons are the electronic components used to do this. What was exciting about this was that it seemed possible to recreate the brain electronically, so that intelligence and thinking could also be controlled by an inanimate electronic structure.
So I began to search for the brain's circuit diagram. It was a good idea to start with the brains of simple animals and then work my way through. At some point I realised that there was something like a common thread in the brain development of the individual species. Gerhard Roth, one of Germany's most important brain researchers, has described this in his book "Wie einzigartig ist der Mensch? The long evolution of the brain and mind". He encouraged me to develop my own brain theory that took into account the evolution of nervous systems and was able to explain which neuronal substrates in today's brains bear witness to the gradual development of brains in the course of evolution.
When I had already written my first two monographs and posted them on the Internet because no publisher was willing to publish them at a loss, I asked myself about the organisational principles to which the brain is subject. What laws, for example, led to the division of vertebrates into reptiles, birds and mammals?
A long time ago, I suspected that the brain must somehow be organised in double layers. Double layers because there appears to be a dualism of on-signalling and off-signalling in the vertebrate nervous system. Receptors translate the strength of a primal magnitude into a firing rate. If the firing rate also increases as the strength of the original size increases, then we speak of an on-signal. However, if the firing rate decreases as the signal strength of the original size increases, then we have an off-signal. Mathematicians speak here of strictly monotonically increasing and strictly monotonically decreasing functions.
Both versions exist in the visual field. For example, there are ganglion cells in the retina that fire more strongly with increasing brightness, but also those in which the firing rate decreases with increasing brightness. For the colour receptors, there are also such on and off variants in the retina. As a result, the visual thalamus, the corpus geniculatum laterale, is a layered structure because the neurones for on-signals form their own independent neurone layers, which are clearly separated from the layers belonging to the off-signals.
The layered structure of the human visual thalamus has already been very well researched. Just think of David H. Hubel and his findings. The human visual thalamus is a layered structure. So is the cortex, but also the optic tectum and the semicircular torus.
In birds, this layer formation is missing and is referred to as nucleisation.
But does this reflect reality? Is it not rather the case that the bird brain could have developed from the reptilian brain and evolved into the mammalian brain? Then the layer formation would have already been present at the beginning. Has it been lost in the course of development or has it been preserved and retained in some way?
So are there three time levels in brain development, in which the first, early time level can be assigned to the reptilian brain, the second, later time level is assigned to birds and a final, third time level corresponds to mammals? Then differences would have to have developed that still exist today!
It was therefore necessary to clarify why the stratification in the vertebrate series occurred and what happened to it.
The starting point for the explanation is the separation of modalities in the vertebrate brain. In the motor area, each joint is generally divided into extensor and flexor muscles, which always act against each other. While the flexor muscles reduce the joint angle through their contraction, a contraction of the extensor muscles causes an increase in the joint angle. As both types act against each other, they represent independent modalities in the motor area. They could certainly be described as on-off variants, as they work against each other in principle.
With the development of the tendency for neurons to separate their axons spatially when they belong to different modalities, such a separation also took place in motor function. As a result, a spatial separation of axons can be observed in the human spinal cord. The axons of the extensor muscles form independent axon bundles, as do the axons of the flexor muscles. The motor on-signals of the flexor muscles take different, separate paths and separate themselves from the motor off-signals of the extensor muscles.
However, not only did the axons of different modalities separate, the associated neurons also took part in the separation. As there are only the two modalities of extensor and flexor muscles in the motor area, the easiest way to achieve the spatial separation of the neurones was to form neuronal double layers. One neuron layer then contains the neurons belonging to the flexor muscles, the second neuron layer contains the neurons belonging to the extensor muscles.
Since each flexor muscle is in principle associated with an extensor muscle, the neuronal double layer can be easily organised topologically, for example by arranging the neuron of the flexor muscle at the bottom and the neuron of the extensor muscle exactly above it, if the layers are arranged horizontally.
These double layers now theoretically occur in all neuron nuclei that receive the muscle tension signals from the signalling motor receptors.
Naturally, the question arises as to when this double layer formation in motor skills could have occurred.
There is a simple answer here. As long as we are talking about the trunk joints of the simplest vertebrates, the flexor muscle of a vertebral joint was located on one side of the body, while the extensor muscle was located on the other side of the body. The extensor and flexor muscles were therefore located on opposite sides of the body. If one follows the theorem of the separation of the modalities, then the axons of the involved extensor muscles were located on one side of the body and those of the flexor muscles on the other. The separation of the modalities was therefore achieved by the fact that the nervous system was also organised bilaterally. One half represented one side of the body, even if for certain reasons there was a signalling junction in which ascending and descending signals changed sides of the body.
However, it is important to remember that all motor receptor signals in the early knitting ladder nervous system travelled up the head. They were passed on from segment to segment on the sensory side. This is how they finally reached the uppermost segment, which we generally refer to as the cortex in our monographs.
Now a contralateral inhibition was realised in the early knitting ladder nervous system. Both halves of the body were in neuronal competition with each other. This was advantageous because it allowed the stronger signals from one half of the body to prevail.
In the top floor, the uppermost segment, i.e. the cortex, the contralateral inhibition was realised by the class 2 neurons present on the sensory side (the second neuron layer from the top) receiving the ascending signals and pulling them to the opposite side with their axons. This is because the nervous system was also bilateral and had two halves.
Once on the opposite side, the class 2 axons were able to activate inhibitory interneurones, which in turn inhibited the contralaterally associated class 5 motor neurones. This is how the contralateral inhibition in the cortex segment was realised in the early days.
But evolution continued. With the development of the basal ganglia, the inhibitory interneurones were lost from the cortical segment. They were now found in the striatum. As a result, the class 2 axons contacted newly formed class 5 neurons on the contralateral side, turning the motor cortex in this area into a double layer as well. One neuron layer represented the motor signals of one side of the body, the second layer the signals of the contralateral side. Originally there was only one such layer.
The new signals from the new layer now also travelled down to the nucleus ruber. But they were on the wrong side of the body, their motor targets were contralateral. They therefore had to switch to the other side of the body. To do this, they used the side-switch nucleus of this floor.
They travelled to the olivary nucleus. There was already an on-layer of input neurons that received the excitatory signals from the opposite side. The new axons with the contralateral off-signals generated new olivary neurons that formed a second layer. The first neuron layer represented the on-signals of the muscles of the opposite side. The off-layer, on the other hand, represented the off-signals from the same side of the body, but which had travelled to the other side via the change of side in the cortex and were able to return to the original side via the olivary nucleus.
For each muscle on one side of the body, there was now a neuron in one of the two layers in the olivary nucleus, and the corresponding contralateral muscle sent its signal to the associated neuron in the second layer. This double layer of neurones was thus topologically well organised.
These double-layer signals travelled from the olivary nucleus into the spinocerebellum. This is where the first problem arose. The spinocerebellum was generally three-layered in structure and had only one input layer for the climbing fibre signals from the olivary nucleus.
Therefore, the double layer signals in the cerebellum had to dissolve. The on-off bilayer split alternately in the cerebellum. In each case, an on neuron of the lower bilayer came right next to an off neuron of the upper bilayer. Like a chequerboard pattern of white and black squares, the Purkinje cells with the motor on-signals of one half of the body alternated with the off-signals of the other half of the body in the cerebellum cortex.
We also find such an alternating arrangement in the visual cortex. While in the thalamus there is still a stratification according to ipsilateral and contralateral signals, an alternating arrangement is observed in the visual cortex. One neuron column receives input from the ipsilateral side, and another neuron column is located exactly next to it, which receives the contralateral signals.
This is exactly how we have to imagine the development in the spinocerebellum. With the one difference that the input did not come directly from the receptors, but from the bilayers in the olivary nucleus. In each case, one Purkinje cell received the ipsilateral signal, while the neighbouring Purkinje cell received the corresponding contralateral signal.
The output of the spinocerebellum travelled to the nucleus ruber and from there via the tractus rubrospinalis to the trunk muscles.
However, since after the formation of the bilayers in the olivary nucleus
and the alternating splitting in the cerebellum, part of the signals represented the opposite half of the body, the rubrospinal tract had to split into a part that supplied one half of the body and a second part that switched to the other side of the body. However, it should be noted that the signals from the trunk muscles only form a small part of the overall projection of the rubrospinal tract, as the signals from the extremities are also included. However, the extremities were only just emerging at this stage of development, and fish were the first to produce them in the form of fins.
At some point in the evolution of vertebrates, the extremities developed. These were often bilaterally symmetrical. This applies to most fish fins, but also to the later tetrapods.
With the beginning of the development of the extremities, it was the first time that the extensor and flexor muscles of a joint were located on the same side of the body. If the signal of the flexor muscle is described as an on-signal, the signal of the associated extensor muscle was an off-signal and its firing rate was the reverse of the firing rate of the flexor muscle.
If one follows the theorem of separation of modalities, the neurones involved in the further processing of the motor signals of such a joint could also form double layers.
The development of these bilayers was accompanied by the development of corresponding markers. In the nervous system, the topology is (in my opinion) largely guaranteed by markers. If the receptors of the flexor muscles produced one or more types of markers, while the receptors of the extensor muscles produced a different type of marker, then these marker substances could be involved in the formation and organisation of the bilayers. We will see that these differences in the marker substances of neuronal bilayers can still be detected experimentally today. The reason for this is the transmission of neuronal markers via the axons of the neurons concerned. Therefore, such markers can often also be detected in the target structures. In the cerebellum, this has already been demonstrated experimentally, as mentioned above.
This separation of modalities in the motor area will certainly have developed gradually, but it is likely to have prevailed in the end in view of the advantages.
The axons of the tendon organs of the limb muscles therefore travelled to the target structures involved and now docked onto the associated neuronal bilayers, controlled by the on-off marker substances.
We simply name the stations of the projection paths involved:
- motor input nuclei of the input floor of the brain (nucleus cuneatus and gracilis)
- sensory thalamus
- sensory cortex
- motor cortex
- nucleus ruber, from there to the spinal cord and to the
- Nucleus olivaris
- Spinocerebellum of the opposite side
- Cerebellar nuclei.
The output of the associated cerebellar nuclei will be discussed later.
We begin our consideration of the consequences of the double layers in the olivary nucleus. According to the double layer theory, the olivary nucleus will form a double layer of neurons in relation to motor limb signals. One layer receives the signals from the flexor muscles of a limb joint, the second neuron layer receives the signals from the extensor muscles. The topology is strictly adhered to. If you imagine a section of the olivary nucleus arranged horizontally and think of the neurones of the extensor muscles at the bottom, the input neurone of a flexor muscle is located exactly above the input neurone of the extensor muscle.
This topology represents the starting system that we can assume for fish, for example, but perhaps also for reptiles.
We can assume that all other intermediate stations of the motor signals were also organised as such double layers.
Part of this double layer arrangement is the fact that each of the two layers had its own neuronal marker responsible for maintaining the topology.
However, nervous systems are subject to evolutionary changes. One major change was the emergence of signalling divergence. Neurones can die. It was therefore favourable if signals were divided among several neurons involved, so that a signal could be transmitted even if a single neuron died. This is a major reason for the reliability and robustness of signal transmission in higher nervous systems.
We assume that such a revolutionary change did not begin in evolutionarily ancient structures, as these were very conservative and their genetic programmes were practically unchangeable and very robust. Therefore, the signalling divergence could only begin in an evolutionarily younger structure.
The evolutionarily youngest new formation in the vertebrate brain was the cerebellum with the involved nucleus olivaris. We therefore postulate that the signalling divergence began in the nucleus olivaris and continued in the cerebellum for the reasons mentioned above.
What does this mean?
Previously, there were two layers of input neurons in the olivary nucleus and, consequently, two layers of output neurons with their own neuronal layer markers.
If additional output neurones were now formed, the two input signals per simple joint had to be divided between several output neurones. Interneurons, which generally existed in all neuronal nuclei and which transferred the input to the output neurons, could be helpful here.
If the interneurons made no distinction between the two layers, they could receive input from both layers and pass it on to the output neurons.
After the formation of signal divergence, there were two input signals per simple joint in the olivary nucleus and therefore also two input neurones, one with the on signal and one with the off signal. If these are divided into several, for example output signals with the help of signal divergence, there were output neurones per simple joint. This is shown in the figure below.
Below you can see 5 input neurones (red) that belong to 5 simple joints and receive the on-signal from the muscle in the olivary nucleus. The corresponding off-signals arrive in 5 input neurones (green) of the lower input layer. Before the signal divergence, there was only a single double layer. The development of signal divergence created the output layer of the olivary nucleus, which is highlighted in yellow in the figure. As an example, 6 output neurones are shown per joint (yellow), each of which receives input from both the on neurone and the off neurone of the associated joint. It adds these two signals together. For each joint there is a vertical neuron column consisting of the on-input neuron, the (here 6) output neurons and the off-input neuron. The output of this circuit leaves the olivary nucleus and is transmitted to the cerebellum via the climbing fibre projection, thus forming the climbing fibre input.
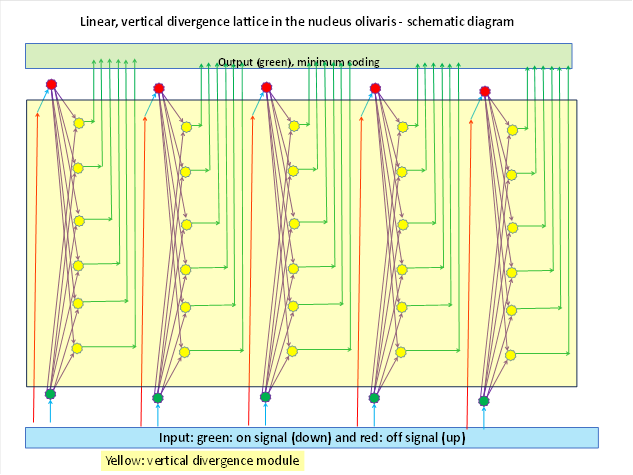
The neuronal marker substances were also passed on to the output neurons via the interneurons at the same time as the signals were passed on, so that the two marker types for on and off signals could also be mixed.
We supplement the figure above by showing the marker substances in colour.
We label the marker of the on-signals red, the marker of the off-signal green . These marker substances are already present in the input neurons and are passed on to the output neurons of each cell column via their axons. Here we assume a decrease in the concentration of the marker substances with increasing distance from the input neuron. Therefore, there is a gradient for each of the two markers. In the upper area, the on marker shown in green predominates and its concentration decreases towards the top. In the lower area, the off marker shown in red predominates and its concentration decreases towards the bottom.
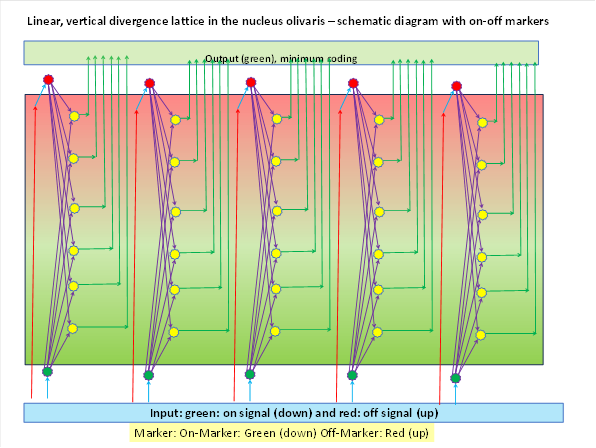
We assume that this marker concentration is passed on to the cerebellum via the output axons, i.e. the climbing fibres. It should also be detectable there.
As the signal divergence distributes not only the input signals but also the neuronal input markers to the output neurons and their concentration decreases with distance, there are two marker gradients within the output layer. The concentration of the neuronal marker of the extensor muscles (off-marker) is highest at the top and decreases downwards until it reaches zero. At the bottom, the concentration of the on-marker of the flexor muscles is strongest and decreases upwards until it reaches zero. This gradient distribution is passed on to the Purkinje cells of the spinocerebellum via the climbing fibres of the nucleus olivaris.
However, the cell layer of the Purkinje cells is single-layered. Therefore, each cell column is folded from the vertical position to the horizontal position at each joint. While in the nucleus olivaris the cell columns are aligned vertically to the surface of the neuron layer, in the nucleus olivaris they form horizontal Purkinje cell rows that always begin with the on-signal and end with the off-signal. In between are the Purkinje cells, in which mixed signal components are present, but within this horizontal cell row of Purkinje cells the neuronal gradients of the ON-Off markers from the olivary nucleus can be found with topological precision.
If the first Purkinje cell on the left of a joint has the on marker, which belongs to the flexor muscle, the marker concentration decreases steadily towards the right until it has a value of zero in the last Purkinje cell of this neuron row. Conversely, the right, last Purkinje cell of a joint has the highest concentration of the off-marker, which belongs to the extensor muscle. This Purkinje cell row forms a kind of circular arc in the spinocerebellum. The neighbouring joint of the same segment on the same half of the body formed another arc of Purkinje cells, which was arranged exactly next to it. Thus, each arc corresponded to a joint of the respective segment of one half of the body.
The rows of Purkinje cells behind them, which were also arranged in such arches, corresponded to the joint muscles of the neighbouring segments.
The arches of Purkinje cells are clearly recognisable in the section of the cerebellum.
If we now look at the on-off markers in such a chain of neighbouring arcs of Purkinje rows, each of the two markers forms a stripe pattern. The ON marker and the off marker alternate in these chains of Purkinje cells because each joint in the nucleus olivaris corresponds to a vertical cell column that already has the gradient structure of the markers and has only been folded over into the horizontal position in the cerebellum.
This is shown in the following illustration. Both the nucleus olivaris and the cerebellum can be seen. The cerebellum surface can be seen here from above. The Purkinje cells are only shown symbolically as small blue circles.
Each climbing fibre arriving in the cerebellum from the nucleus olivaris contacts a group of Purkinje cells in the cerebellum (4 in the figure below) and runs from bottom to top in the figure. In contrast, the mossy fibres not shown and the parallel fibres would run horizontally in this figure. In this respect, parallel fibres and mossy fibres or parallel fibres cross orthogonally (at right angles).
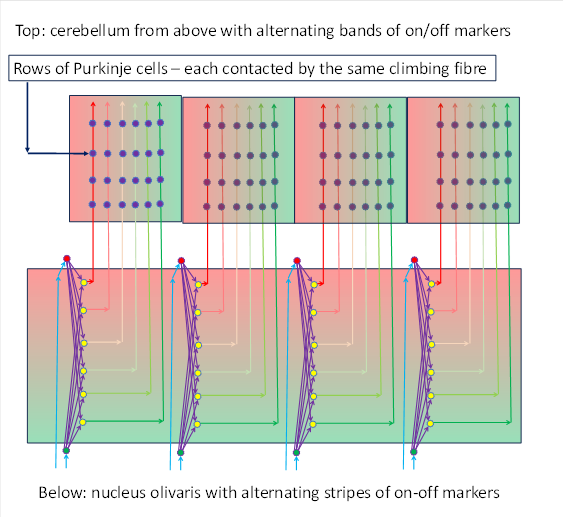
The researchers Joy Zhou, Amanda M Brown, Elizabeth P Lackey, Marife Arancillo, Tao Lin and Roy V Sillitoe succeeded in experimentally demonstrating this alternating stripe structure of two marker types in the spinocerebellum, which they published under the title "Purkinje cell neurotransmission patterns cerebellar basket cells into zonal modules defined by distinct pinceau sizes".
The vertical structure in the olivary nucleus is transferred into a horizontal structure in the spinocerebellum. It should therefore also be possible to detect these marker structures in the olivary nucleus, although they must have a gradient in the vertical direction. This proof is still pending. All brain researchers are called upon to provide this evidence experimentally and to prove the vertical orientation of the signalling divergence in the olivary nucleus.
It should be borne in mind that interneurons themselves do not usually form their own action potentials, but only transmit the neuronal excitation subliminally. As interneurons, they had no myelin and were part of the grey matter. However, this results in signal attenuation during signal transmission. The cable equation for myelin-free axons applies to this type of signal transmission, which states that an exponential attenuation occurs that increases with the distance between the input neuron and the output neuron. This transfer function is a strictly convex function. The superposition of several such functions in turn results in a strictly convex function that always has a global minimum. The position of the minimum results from the signal ratio of the ON signal to the Off signal. It should be noted that the on-signal increases with increasing muscle tension of the flexor, while the corresponding off-signal of the extensor muscle behaves in the opposite direction. This means that there is a clearly defined excitation minimum for each joint angle.
This type of signalling divergence was described in chapter 4.2 "The second expansion phase of the primordial spinocerebellum" in my monograph "Brain theory of vertebrates" (link: https://www.andreas-malczan.de/Gehirntheorie_4_2.html).
The signalling divergence in the nucleus olivaris leads to the minimum coding of the joint angle of the joint involved. After the signal divergence occurs, we can observe that (in relation to the horizontally imagined neuron layer) vertical neuron columns are formed, each of which is assigned to exactly one simple joint.
Within this joint column of neurones, the excitation minimum moves back and forth, i.e. upwards or downwards, when the joint angle changes. At a medium joint angle, the excitation minimum is located approximately in the centre of the neuron column. At the smallest joint angle (flexor muscle maximally contracted) the excitation minimum is at the top, at the largest joint angle at the bottom. This is due to our decision to assign the lower layer to the flexor muscle (which is unfortunately arbitrary and could also be the other way round).
Later, a further divergence of signals was to lead to one joint being associated with several, later even quite a few, such arcs of Purkinje cell rows arranged one behind the other, which gradually led to the transition to the pontocerebellum.
The output of the spinocerebellum was originally motor in nature, travelling from the cerebellar nuclei to the contralateral nucleus ruber, which at this stage of evolution was also organised as a bilayer. The signal axons, which were greatly increased in number due to signalling divergence, formed the magnocellular part in the nucleus ruber, in which the signalling divergence was reversed by a corresponding signalling convergence, so that the original signals of the joint could reach their motor targets, the muscles.
However, in the course of evolution, an ascending projection also became established and the output of the cerebellum found its way upside down.
However, this led to different solutions, which led to the splitting of the vertebrates. At the latest when the vertebrates had transformed their previous fins into extremities and left the water, they split into reptiles, birds and mammals.
The reason for this was the different path taken by the ascending cerebellum signals.
In reptiles, these signals formed a new level in the previously segmented rope-ladder nervous system. They ascended on the sensory side, but always remained close to the ventricular cavity as new signals. New neurones were always formed on the inside of the neural tube, while the axons ran on the outside. When the new cerebellum axons had reached the upper ventricular space of the head area, they remained inside and travelled in the ventricular space quasi below the pallium to the motor side, from where they descended again to the nucleus ruber. Due to the divergence in the olivary nucleus, their number was now considerably increased. However, as there was enough space in the height, the neurones belonging to a joint were able to orientate themselves vertically to each other again and thus adopt exactly the orientation that they originally had in the olivary nucleus. This meant that the sensory side of this neurone structure in the ventricular cavity was orientated vertically. Each joint corresponded to a vertically arranged neurone column.
The signals were now passed on to class 3 neurones - as is usual in the rope ladder nervous system - which travelled to the motor side. There they were able to descend to the substantia nigra pars compacta and find their way into the basal ganglia system. They also found an ascending path into the limbic system.
On the other hand, it proved expedient to reverse the signalling divergence of the olivary nucleus. Therefore, these signals on the motor side converged on class 5 neurons to restore the original joint signals. As a result, the motor side of this structure was a vertically orientated convergence system. This structure, located in the ventricular cavity, is called the dorsal ventricular ridge (DVR). It is a conspicuous structure in reptiles.
In birds, this structure is also still partially present, but it is also found on the outside of the brain. Apparently, the ascending axons of the spinocerebellum also found their way to the outside of the brain and formed a new structure there, known as the hyperpallium. The motor part of this receives the motor cerebellum output and is also vertically orientated. In its organisational form, the (motor) hyperpallium thus resembles the nucleus olivaris, which contains vertical joint columns and has a vertical gradient orientation of two complementary types of marker substances. Precisely these vertical marker gradients should also be detectable in the hyperpallium and DVR. Evidence here could prove the correctness of my theory!
By using the hyperpallium to analyse signals, birds were able to achieve higher intellectual performance than reptiles, in which the ventricular cavity could only provide limited space for the DVR.
This vertically orientated structure was also formed in mammals, but in mammals it is referred to as the frontal cortex. However, this structure was able to develop further in mammals through evolution, so that we no longer find the original state that corresponds to the DVR or the subpallium. In addition to the old, vertical layering in cell columns, a signalling divergence also occurred in the frontal cortex in the lateral plane. Additional output neurons formed between the cell columns of the different joints, which now received signals from different, neighbouring joints. And the signal divergence in the surface led to these new output neurones being besieged by countless interneurones, whose signals they picked up. However, as the input neurones formed a thick layer, the output neurones were also distributed in this layer. This resulted in a spatial divergence system, as described in chapter 4. As a consequence of this lateral signalling divergence, the frontal cortex increased greatly in area, and the onset of fold formation enabled this larger area to be accommodated in the available space, which also increased. In the non-motor cortex areas of mammals, there was also a signalling divergence in area, whereas in birds there was only a vertical signalling divergence on the sensory side and a vertical signalling convergence on the motor side.
This enabled the mammals to analyse joints with several degrees of freedom using signal technology and, for example, to analyse the arm rotation of a ball joint in space.
Mammals are therefore masters of multidimensional signalling divergence. As this is also mastered by them in non-motor modalities, this resulted in a tremendous gain in information, which ultimately, in conjunction with the developing pontocerebellum, made higher intelligence possible. This will be explained in the next chapter.
It must be remembered at this point that the neurons in the nucleus olivaris belonging to a joint column represent a minimum-coded signal vector. The signal inversion in the spinocerebellum results in a maximum-coded signal vector for each joint. This maximum-coded signal vector is also found in the DVR of reptiles and birds, in the hyperpallium of birds and in the frontal cortex of mammals as a result of signal propagation. Only these maximum-coded signal vectors are really useful for the motor control of the muscles.
Monografie von Dr. rer. nat. Andreas Heinrich Malczan