Human brain theory
for the European Union’s Human Brain Project
ISBN 978-3-00-068559-0
2 The splitting of modalities and module formation - prerequisite for the division of labour in the brain
The splitting of the modalities began gradually and took place step by step. Different receptors delivered signals of different modalities. Already when the signals arrived in the neural tube, where they ended at neurons of class 3 and 4, the separation occurred. Different receptor types occupied their own neuronal layers in the neural tube, assigned only to them. Here, the spatial cohesion of neurons of one modality was so strong that it was cross-segmental. Neurons of the same modality came together in one layer in the neural tube even when signals from different segments corresponded to them. Then they were spatially adjacent within the layer if the segments were adjacent.
In the process, a topological well-order was formed. Signals from neighbouring receptors of the same modality ended in the neural tube in neighbouring neurons. Since sensory signals - and all receptor signals were sensory in nature - travelled headward from all segments, each head segment had a complete copy of all receptor signals from the trunk segments. Here, the arrangement of the receptors in the body surface (skin) or in the body was transferred from the receptors to the input neurons of the body model in the respective head segment.
Thus, each head segment had a complete body model of the trunk receptors, which was topology-preserving.
The topmost head segment corresponds to the cortex in vertebrates. Here, too, the trunk signals ended up topologically well-ordered and reflected the arrangement of the receptors in the animal's body. This topological arrangement is also called the homunculus. However, it is present several times in the cortex. Apparently, the headward axons, which carry the signals of different modalities, separate before reaching the first segment, each modality moving into an independent cortex area.
This is what we call the splitting of the modalities. It once began with the fact that a tripartite division of the receptor types was already formed in the rope ladder nervous system, so that the rope ladder split at the upper two segments. Instead of one ladder, from the second segment onwards there were three ladders, each for a specific modality group.
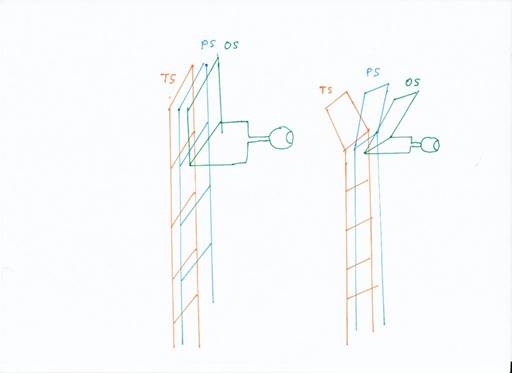
Figure 4: The breakdown of modalities
Since the signals in the sensory spars moved upwards, then reversed direction in a kind of loop and descended again, we also call the top floor a loop (or reversal loop). Then, after the primal modalities split, there were three loops: the temporal loop, the parietal loop and the occipital loop.
These three ladders gave rise to three vertebrate cortex lobes, the parietal lobe, the temporal lobe and the occipital lobe. The parietal lobe received the signals from the receptors directly related to motor function, such as the muscle tension receptors, the receptors for tactile stimuli and pain stimuli. The temporal lobe received the receptor signals from the hair cells of the vestibular system, later the lateral line signals and the auditory signals. The occipital lobe received visual signals.
With the formation of the cerebellum, the frontal lobe emerged, which receives the output of the cerebellum.
These are the four most important lobi of the brain.
During the evolutionary higher development of the vertebrate species, the splitting of the modalities progressed further. Now the modalities separated not only at the level of the second segment, whose sensory and motor nuclei are now called the thalamus, but already in deeper segments.
The separation finally went so far - for example in primates and humans - that certain muscle groups formed their own input nuclei in the head segments, which received their own representation in the cortex. This was an enormous advance for the interaction of signals from different modalities, because now, for example, eye movements, neck movements and movements of visual objects could be coordinated with each other in such a way that a stable image was created, independent of one's own or other people's movements. This was only possible because a clean separation of the modalities had been made beforehand.
The splitting of modalities reached a particularly high degree of development in humans. If new submodalities arose for a modality, a separation of these submodalities into separate layers occurred in the input layers of the second and first floor, i.e. in the thalamus and the cortex.
This separation into submodal layers did not begin with humans, of course, but much earlier, probably already with simple vertebrates. However, it reached such a high degree of development in humans that their brains can perform feats that are either impossible in the animal kingdom or only occur in a few species - and then only selectively as a special case. For example, some bird species can perform cognitive functions that the rest of the animal kingdom is incapable of, but humans can.
The splitting into submodalities is to be explained by way of example.
Originally, there was only one type of receptor for visual perception: the green receptor. It supplied input layer 4 with signals that evaluated the current brightness. The output was transmitted to the neighbouring layers 3 and 2 and 6. Layer 3 projected to the motor centre, layer 2 to the lateral switch nucleus for contralateral inhibition and layer 6 supplied the mean nuclei. In the following figure, this is interpreted as stage 1.
Through gene duplication, a new receptor was formed: the red receptor. Now two colours could be perceived.
This led to a splitting of the fourth input layer in the sensory nucleus of the second segment - the visual thalamus. There, the neurons received the input from the retina. Layer 4 split into the sub-layers S4-green and S4-red. Here - according to the hypothesis - each of the sublayers had its own output layer of class 3 neurons.
Via class 3 neurons, this input moved to the motor part of the visual thalamus and ended there at class 5 neurons, which projected to the motor neurons. This enabled the animal to respond to visual stimuli with movements that caused it to approach food or flee from predators.
Via class 2 neurons, the signals moved to the opposite side to inhibit contralateral visual stimuli there. Thus, both eyes were in neuronal competition, the stronger signal suppressed the weaker one and prevailed. Larger prey on the left side was eaten preferentially if the prey on the right was significantly smaller.
The visual mean nuclei were activated via class 6 neurons and cranked up foraging via back projections to sensory and motor centres.
When the green receptor split and a new red receptor was created, this also led to the splitting of the fourth input layer in the thalamus and cortex.
It disintegrated into two sub-layers. Layer 4, which already received input from the green receptor since time immemorial, remained. However, we give it a new name, it is now the layer 4-green.
The new red receptors also sent their signals to layer 4. Initially, the green and red signals in layer 4 may have been mixed. But like goes with like! So the input neurons that received the red signals began to migrate upwards, and then (after thousands or hundreds of thousands of generations) formed their own layer 4-red. Even today, the view is widespread in neurology that new (evolutionarily younger) neurons always attach to the outside of the existing ones and form their own layer.
This completed the separation of the two submodalities in the visual thalamus. However, in the course of evolution, it also transferred to the superior segment. Thus, the signals ascending from the thalamus via the neurons of class 4-red and 4-green also separated in the fourth cortex layer. The fourth cortex layer split into the 4-green and 4-red sublayers. Again, the evolutionarily newer layer - the 4-red layer - was located on top of the older 4-green layer. In the following figure, this is marked as stage 2.
Still later, when the blue-sensitive receptors emerged in the retina, three types of receptors were available for layer 4 in the thalamus and cortex. Therefore, a new layer 4-blue had to emerge, which was again spatially located at the top because it was the youngest layer.
Originally, layer 4 had layer 3 above it. When the splitting of the modalities began, this layer 3 was also included in the splitting. Each new sublayer 4 received its own sublayer 3. This is already taken into account in the figure below. This probably applied to all six neuron layers, which now formed their own sublayers.
In humans, however - and possibly already in earlier vertebrates - layer 4 blue also (additionally) arranged itself below the previous layers. One reason was (possibly) the signal relationship between the green-sensitive and the blue-sensitive receptor. The latter was more or less the "grandchild". Thus there were two blue layers, one at the top and one at the bottom. This is illustrated in the following figure in stage 3.
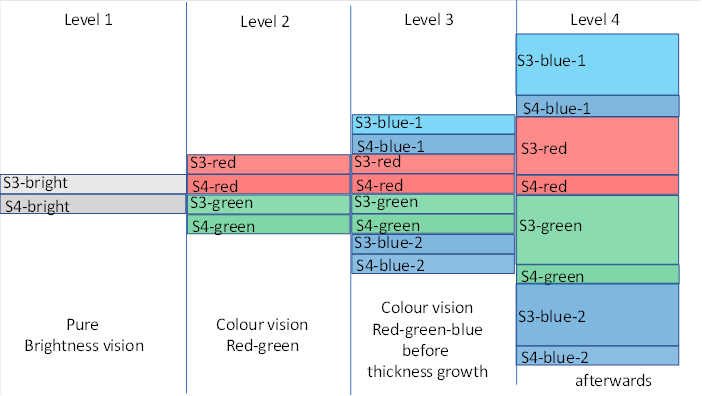
Figure 5: Splitting the visual modalities
The advantage was obvious. Now one could see the single colours green, red and blue as well as mixed colours, such as red + blue = violet, red + green = yellow or green + blue = green-blue. Then the output neurons of each of the two neighbouring layers were active at the same time. We call this additive colour mixing. This now also took place neuronally. A simple calculation example should clarify this.
Let a retinal red receptor excite a ganglion cell whose firing rate reaches an input neuron in the visual cortex at a frequency of 100 action potentials per second. This is located in layer S4-red. Its excitation activates an output neuron in layer S3-red. At the extremely short distance - both layers are directly adjacent - the excitation passes completely to the output neuron, which then also fires at 100 Hz.
If, instead, a green receptor fires just as strongly because green light excites it, it activates an output neuron in the S3-green layer. This now also fires at 100 Hz.
Let us now imagine that both types of receptor are excited because yellow light falls on the retina. However, yellow light excites the red receptor, as can easily be seen in the characteristic curve diagram of the retinal sensitivity curves of the different visual pigments. Somewhat less than red light would. For example, the red ganglion cell only fires at 80 Hz.
At the same time, the green-sensitive ganglion cell fires, because the green visual pigment also reacts to yellow, albeit more weakly than to green. Thus, an output neuron also fires in the S3-green layer at 75 Hz, for example.
Since the red receptor arose from the green receptor, the neurons for the signalling pathway of the red and green colours were signal-related. Therefore, the output neurons of layer S3-red could also receive excitation from input neurons of layer S4-green. Thus, a subgroup of neurons formed there that could receive both types of signals.
Thus it came about - in the course of evolution - that when an input neuron in layer S4-green and in layer S4-red were simultaneously active, a common output neuron in layer S3-green was contacted and also excited. We will provisionally refer to this outout neuron as the addition neuron. In addition to those already excited in layer S3-green and S3-red. However, these only received input from one neuron, firing at 80 Hz for green and 75 Hz for red. Now there was one that received input from both. Its firing rate was greater than 80 and also greater than 75 Hz, for example 155 = 80 + 75.
Thus, there are three output neurons for each image pixel of the retina: One is responsible for the red colour and one for the green colour, the third was supplied additively, i.e. received the input for the red and for the green colour at the same time. If both the red receptor and the green receptor were excited, the output of the additive neuron was significantly stronger than the output of the two neurons for red and green.
This is where neighbour inhibition comes into play. The firing rate of 155 Hz is much stronger than that of 80 Hz or 75 Hz. It inhibits these two firing rates or the neurons that produce them. With total inhibition, there is only one output: f = 155 Hz. And it clearly signals: Yellow light detected.
This is because the rest of the system's output is negligibly weak because it is a victim of the mutual inhibition of the output neurons.
This is how the visual cortex learned to recognise the colour yellow. The same train of thought can be applied to the mixed colours violet = red + blue or blue-green = blue + green. This system could recognise colours that were composed of two of the three primary colours red, green and blue in an additive manner.
It must be pointed out that the splitting of visual modalities is not only detectable in the human cortex. It already occurs in the retina. There, the different receptors are arranged in different layers of the retina. And already in the retina there are neurons - they are called ganglion cells there - which receive input from different layers. This input can be excitatory, but also inhibitory. For example, there is a type of ganglion cell that is excited by the green receptor but inhibited by the red receptor. We call them green-on/red-off ganglion cells. Similarly, there is a type of ganglion cell that is excited by the red receptor but inhibited by the green receptor. They are called green-off/red-on ganglion cells. Another type of ganglion cell is of the blue-on/yellow-off type. Furthermore, there are - in principle as neuron class 6 - large mean neurons, which tap all On ganglion cells and represent the Hell-On type. Likewise, there is the magnocellular dark-on type. In this respect, the retina already represents a module. It already delivers brightness-weighted and colour-weighted signals to the tectum opticum, the visual thalamus and the cortex. The layering in the retina continues in the thalamus and cortex and is further refined.
In humans - according to my theory - the splitting of the modalities in the colour system continued. While in the primary visual cortex the signals of the three different colour receptors still ended in different sub-layers of layer 4, a new subsystem emerged above the previous cortical floor, a kind of new floor of the cord ladder system. There, the colour information no longer ended in three different input layers, but in a single layer. In exchange, however, the signals split up and moved alternately to this new floor. So it is possible that there, in the secondary visual cortex, a new sensory input area was created. In this input area, the retina was again mapped retinotopically. However, each retinal pixel corresponded to three input neurons, which formed a triangle on their side. In one corner of the triangle was the input neuron that received the red-on/green-off signal from the primary cortex. In the second corner of the triangle was the input neuron that received the green-on/red-off signal from the corresponding retinal pixel. In the third corner of the triangle, the blue signal arrived, but was inhibited by the colour yellow and therefore represented the signal Blue-On/Yellow-Off. Thus, each individual retinal pixel was associated with a colour triangle whose corners were assigned the primary colours red, green and blue. In this cortex area, the retina roughly corresponded to the area of a colour television, in which each pixel consisted of three light points to which the primary colours red-green-blue were assigned. Additive colour mixing creates a coloured image of the world for us. And this is exactly how the secondary visual colour field is organised.
This secondary visual colour field also projected (among other things) into the basal ganglia and the limbic system. In addition, a tertiary cortex area developed that received both the input from the secondary colour field and the time-delayed return output of the basal ganglia, so that changes and movements became perceptible there. The reverse output of the limbic system led directly to false colour afterimages in complementary colours.
Such a split is also observed between the thalamus and the primary cortex. In the visual thalamus, the signals from the left and right eyes still end up in different layers, which are, however, well-ordered retinotopically. In the visual cortex, however, these signals are no longer arranged in superimposed layers, but form alternating eye-dominance columns whose input comes alternately from the left and right eyes. Instead of layering, alternating arrangement occurs. And for the colour system, I also postulate a transition from the layer system in the primary visual cortex to the secondary cortex, where the colour signals previously arranged in three layers are now alternately arranged in the surface, thus forming the colour triangles.
However, this development was not the end point. The colour system managed to recognise millions of colours through a small change. However, not all species were able to do this. Excellent colour vision required an important but relatively simple change in the structure of the class 3 outpouch layers: thickness growth and width growth. We discuss here first the thickness growth. This is shown as stage 4 in the diagram above. In my earlier monograph, I theorised that this thickness growth occurred predominantly in reptiles and birds. In reptiles (and birds), this is how the dorsal ventricular ridge (DVR) formed. In birds, this structure also occurs on the surface of the brain, where it is called the hyperpallium. Both structures are apparently vertical divergence modules.
In vertebrates, development may also have begun with the growth of thickness, but - in a number of species - there may also be growth in area, or in primates even growth in thickness and area.
How did the growth in thickness initially occur?
Each outpu layer of neuron class 3 began to form reserve neurons. Neurons could die. If there were spare neurons nearby, they could step in as replacements.
Thus, it became standard (in higher species) that more neurons were formed in the outpu layers of class 3, but also class 2, 5 and 6, than were strictly necessary.
This is where a problem arises for neurologists: neurons with identical signals always inhibit each other when they are active. This leads to atrophy of the supernumerary neurons, so that only one remains.
However, this atrophy does not occur if the neurons differ in activity, i.e. strictly speaking, they do not process identical signals, but only similar ones.
Two output neurons that receive input from one or more neurons form slightly different signals precisely when the pathways of the signals to them differ. If one neuron is located closer to the input neurons while the second is further away, their excitations differ from each other. This is because damping acts in myelin-free layers. The greater the distance, the greater the damping. If it was convenient to transmit the signals via several signal paths, the neurons involved had to differ at least by the signal damping. Because then they had different firing rates. Their signals were only similar, but not identical. Thus, no atrophy took place.
Therefore, the thickness of the outpouch layers grew. Thus, there were enough reserve neurons. They differed slightly in output behaviour, but sufficiently. Because the distance of an output neuron to the upper and to the lower input layer differed from each output neuron that was located above or below it. All output neurons belonging to a retinal point in the visual output layers with greater thickness formed an eye dominance column. They received input from the inferior and inferior layers simultaneously. Each output neuron was supplied simultaneously by both the superior and inferior layers, the synaptic formations for which were already established during the formation and maturation of the outpouch layer.
But how could this system recognise colours, especially mixed colours?
Here we have to do some mathematics and explain the working principle of signal mixing. We refer to such systems here as a divergence module with vertical signal mixing. This is because the excitations from the colour sensors reach the two input layers - one at the bottom and one at the top, and spread out vertically to mix.
The divergence module with vertical signal mixing is described below. It should already be pointed out here that quite a few species of vertebrates were able to expand this divergence module considerably by also using a width growth. Such divergence modules enable the emergence of orientation columns in the brain for visual direction and contour analysis.
After explaining the analytical properties of the visual divergence module with vertical signal mixing for the parvocellular input, the mathematical derivation of its remarkable properties follows.
One more hint should be given here: Neighbour inhibition in such divergence modules only occurs when the output leaves the divergence module. The output of the most diverse sublayers of class 3 neurons leaves the system in several ways. Upwards, class 2 output neurons are activated, which pass their input to the contralateral side. Neighbour inhibition must take place in this layer.
Furthermore, layer 3 projects to the motor side of the visual cortex. This can be recognised by the fact that the sensory input travels via axons to the motor side and activates class 5 neurons there. Neighbour inhibition can now act between this output. The motor side of the visual cortex is often called the secondary cortex.
The neighbour inhibition is thus outsourced from the divergence modules. Therefore, neurologists analysing excitation with a fine probe in field V1 will often have the impression that all its neurons in an eye dominance column are active. In principle this is so, but nevertheless their excitation varies in intensity. Extreme value coding takes place in the outpup layers. The maximally excited neuron evaluates the respective submodality in terms of its signal strength. Its activity evokes what neurologists and philosophers call qualia with regard to an elementary stimulus.
However, the splitting of modalities went further. If there was a higher segment, signals that had previously reached different layers of a module were arranged side by side in one level in the segment above, whereby the existing submodalities naturally had to mix. However, if there was no higher segment, these signals created a new segment in which this final splitting step was completed. Therefore, above the primary visual cortex, which still arranges its signals sorted for each submodality in a separate layer, there is a higher-level visual cortex (second degree), in which, for example, the colour information of the three primary colours red, green and blue are arranged in a single layer, so that small colour triangles are practically created there. In the course of the signal divergence in this region, the colour triangles expanded and the output neurons of these triangles could in turn carry out a maximum coding of the mixed colour, which was realised by superimposing the excitations of the three input neurons of each colour triangle. Only then could a further and much finer colour analysis be carried out.
This further splitting of submodalities progressed in the motor area as well as in the visual. All modalities are likely to be affected.
Monografie von Dr. rer. nat. Andreas Heinrich Malczan