Human brain theory
for the European Union’s Human Brain Project
ISBN 978-3-00-068559-0
6.1 Convergence modules - inverse divergence modules
For this chapter, we need to activate our insights into brain development from the early double-stranded ladder system of segmented bilateria. There, the sensory signals rose headwards and additionally bent sideways to the motor part on each floor. This was also the case in the first segment, which became the cortex.
Every sensory signal had to move from the sensory cortex side to the motor cortex side. And since the cortex had by now broken down into modality lobes and modality strips in each gyrus, each modality was faced with this task. It had to move to the motor side so that its signals would drive the associated motor neurons.
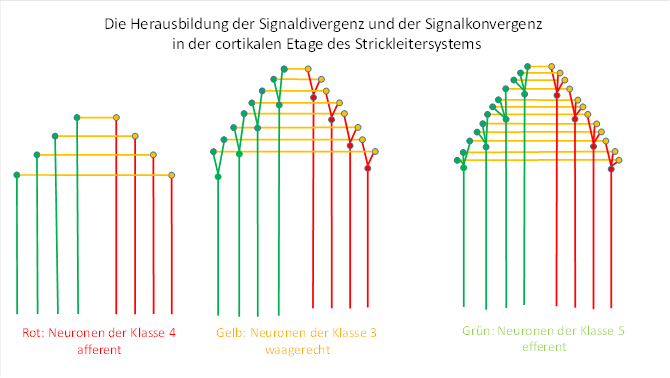
Figure 38: Divergence and convergence in the cortical floor
For example, each modality could use the signals from its receptors to cause movement towards prey, but also flight from predators.
With the emergence of signal divergence on the sensory side of the cortical floor of the early cord ladder system, the ascending signals were divergently distributed among the class 3 neurons. This certainly took place gradually. Initially, a class 4 neuron supplied exactly one class 3 neuron, whose axon moved to the motor side and terminated at exactly one class 5 neuron. Later, the ascending signal was distributed to 2 neurons that moved to the motor side. However, they had only one target neuron on the motor side, to which they both docked together. When a third class 3 neuron was included, three axons arrived on the motor side and contacted the common target neuron. This is shown in the figure above.
In the beginning, the signal divergence certainly served exclusively to increase the reliability. Only later - due to the distance between the input and output neurons - did an extreme value coding of the output occur as a result of the non-linear signal attenuation, which was to the animals' advantage.
The further this development progressed, the greater the number of class 3 neurons became. While on the sensory side the various divergence modules emerged through signal divergence and enabled a much finer analysis of the signals, a corresponding signal convergence had to be developed on the motor side. Thus, the convergence modules emerged on the motor side. They reversed the sensory signal divergence.
On the motor side, a signal convergence took place that completely reversed the signal divergence of the sensory side. Thus, no matter in which segment and on which side of the body the connected motor neuron was located, it received the input it had received before the signal divergence was formed. The motor responses to the sensory signal were preserved.
We found that signal divergence created new submodalities: Brightness, colour, orientation angle of line elements. Movement, etc. The original modality thus disintegrated into new submodalities.
Whenever a modality broke down into new submodalities through signal divergence in divergence modules, the new signals were also sent to the motor part of the associated cortex. It was precisely the cortex region to which the original primordial signals were sent by the axons of the output neurons.
If a signal divergence now took place in the layer of sensory signals, the diverged signals moved exactly into this area and converged again on the original output neurons, which had previously received exactly the original signals.
In the case of signal divergence of motorically effective signals, these moved from the sensory side - where they arrived as muscle tension signals, for example - to the motor side and converged on the Beetz cells of the motor cortex. Their large dendrite trees received the previously diverged signals and combined them again. Thus, the primordial signal on the motor side was created by a strong signal convergence. This reversed the signal divergence.
Now, if a divergence module led to a thickness growth or to an area growth or (mostly) to both, exactly this change took place not only on the sensory side, but to the same extent on the motor side. In the target areas of a module, the signals met an analogously constructed motor module in exactly the same way, which reversed the signal divergence.
For example, if a cube-shaped red/green colour module with spatial signal propagation received the retinal input from a total of eight ganglion cells and generated, for example, 1000 different output signals (10 neurons in width, 10 in length and 10 in height - chapter 1.1.5), it consisted of 10 layers, one above the other, each of which had a square structure and contained 10 neurons in width and 10 in depth.
These neurons projected (among others) to the motor side, which then also consisted of 10 layers of neurons. There they contacted 1000 neurons in a cube consisting of 10 layers of neurons of 100 neurons each arranged in a square of 10 by 10 neurons. But both cubes, the sensory and the motor, were arranged as mirror images of each other. The plane that ran vertically between the cubes was the mirror plane.
Now the signals from the class 3 neurons could move to the motor part and contact the 1000 interneurons in layer 5 without their axons getting in the way and creating disorder. They simply pulled down on the sensory side until they left the cortex layer. Then they moved upwards on semi-circular pathways to the opposite side into the motor area. From there, they continued vertically until each axon had reached the height at which the relevant target neuron of layer 5 was located.
The axons of the class 3 sensory neurons excited these interneurons, whose axons transmitted the signals to the cube corners. Thus, in the convergence module, there were again the eight signals that had previously fed the colour module. They could be received by the output neurons of class 5, which were located in the eight cube corners, and were now delivered to the original signal receivers - the motor neurons.
These convergence modules - also called inverse divergence modules by me - produced the signals of the old modalities again from the signals of the new modalities. However, this assertion must be proven mathematically.
Here we should pursue the thesis that the stratification of the cortex was preserved. Only through this could an inverse module undo the work of the previous analysis module.
Of course, the primordial signals could have gone directly to the motor side. But since the primal signals came from receptors, the receptors could have directly controlled the muscles instead. A brain would then not have been necessary at all. Each receptor could have simply controlled a motor neuron - as in primeval times.
Therefore, the signal analysis in the cortical divergence modules must have had a higher utility, otherwise it would not have arisen.
The first benefit arose in prehistoric times when living creatures acquired the ability to recognise many colours. This facilitated the recognition of food. While herbivores benefited from green-sensitive visual pigments because the green chlorophyll revealed plants, carnivores could reliably detect the red colour of blood. When some species could detect both the colour green and red and also recognised their intermediate colours, omnivores had an advantage. Whether this simple approach stands up to critical analysis is questionable. But hypothesising is the business of researchers. Every hypothesis can - if it does not prove itself - be thrown overboard.
.1 The convergence module for the colour module with vertical signal mixing
We will now examine the operation of a convergence module with vertical signal mixing, also called a vertical convergence module for short. We can also call it an inverse divergence module, mainly because it refers to the way it works. Its input comes from a divergence module. Via axons of class 3 neurons, this module sends its output to the motor side in the convergence module. In both the divergence module and the convergence module, the excitation spreads (primarily) in a vertical direction. The reason is the signal relationship of the input signals of a retinal pixel.
The retina is mapped into the x-y plane. Two points are assigned to each retina point PR(x,y) in the module. The red-on signal with the firing rate f1 reaches the module at the point PM(x,y,1) at the height z = 1. The corresponding green-on signal arrives at the module point PM(x,y,0), i.e. at the height z = 0. Since both signals are signal-related, all neurons with the coordinates y and x receive these two input excitations, regardless of the height z at which they are located. By superimposing both excitations, an excitation maximum is created in the vertical neuron column, the height of which encodes the colour, i.e. also the (average) frequency in the red-green range. The reason for this is the non-linear transfer function, which is also strictly concave. This guarantees a global excitation maximum.
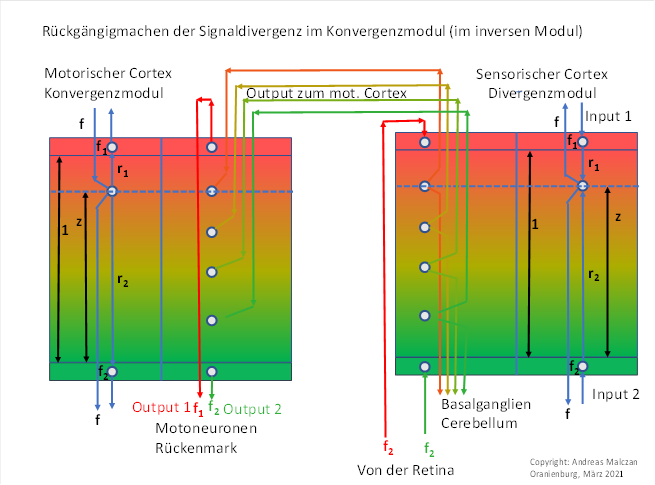
Figure 39: Convergence module for the colour module red-green with vertical signal propagation
Each of the two modules in the above figures is divided into two parts. On the outside, the principle circuit diagram for three participating neurons is shown, the quantities h, r1 and r2 are explained by black arrows and the thickness of the outpouch layer is marked with 1.
As an example, we choose the colour module with vertical spread, i.e. without orientation columns. It serves purely for colour recognition. Its output travels via the axons of the class 3 neurons to the motor side in the associated convergence module. All colour signals of an eye dominance column move to the motor side in a well-ordered manner and (presumably) contact one output neuron per colour signal in layer 5 - the input layer. In the process, the input neurons belonging to a mixed colour arrange themselves exactly in the same order from top to bottom as they were arranged in the sensory divergence module. The colour layering is therefore preserved.
We know that the output of the divergence module is maximum coded. Only one neuron in the entire vertical column is maximally excited. It inhibits the other output neurons of the output column via neighbour inhibition. Thus, of the many colour conductions to the motor cortex, only one is active at a time.
These colour lines - i.e. the axons of the class 3 neurons of the divergence module - run, in deviation from the above sketch, below the cortex layer on long axons well insulated with myelin to the motor cortex. Here, these axons form roughly semicircular connections, with the lower half of a semicircle describing the path of the axons.
We remember: Only one colour neuron at a height z above the x-y plane in the neuronal column of the divergence module is maximally excited. Only its output reaches the motor side, the other colour signals become a victim of neighbour inhibition.
In the convergence module, this colour signal hits an input neuron, which also has height z from the x-y plane, and excites it with firing rate f. Its excitation now spreads upwards and downwards in the module. A thin, single-layer neuron layer of output neurons is located at the upper boundary surface of the input layer, and likewise at the lower boundary surface. The two output neurons of the module are located there, while the input neurons again run like a string of pearls from top to bottom.
During propagation, the distance-dependent damping takes effect. The upper output neuron - it represents the colour red - has the distance r1 from the input neuron. We have normalised the thickness of the input layer to the value 1 (a different layer thickness would not change anything about the basic functioning). The upper output neuron has the distance r1 to the input neuron, the lower one the distance r2. The input firing rate has the value fD.
Thus, for the firing rates f1 and f2 of the two output neurons, the following equation applies
![]()
![]()
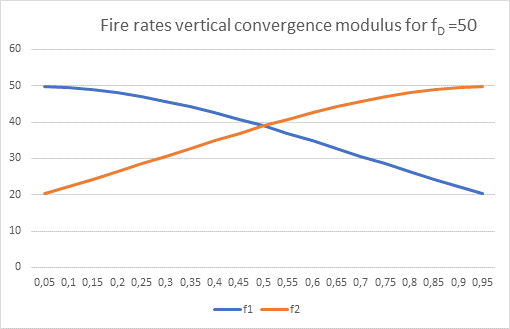
Figure 40: Fire rates vertical convergence module
In the divergence module, the height z of the output neuron changes with the maximum firing rate as a function of the two input firing rates f1D and f2D. This is shown in the figure below.
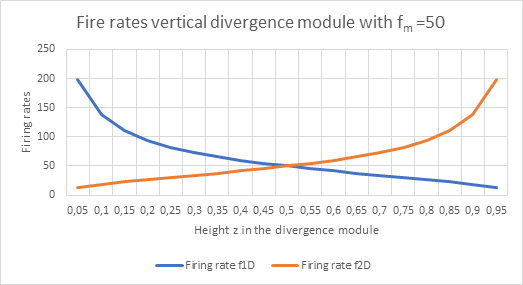
Figure 41: Fire rates vertical divergence module
In comparison, one can see that the convergence module does not exactly recreate the original firing rates that occur as input to the divergence module, but only approximates them. However, this error is certainly much more favourable than the alternative in which no output at all would move from the sensory to the motor side. In this respect, the development of convergence modules was important for the animal's further, mostly motor responses to sensory input.
But it was not until the development of memory modules that the fine analysis of all modalities and the generation of new submodalities through divergence module brought real advantages. Thus, the mere idea of a prey - i.e. the activation of a stored image of the prey - could trigger exactly the reactions that a real prey caused. In humans, images of stored states and signal combinations can be sent to the cortex in this way and trigger all kinds of activities there - from dreaming to searching for food, water or even sex.
Therefore, chapter 7 deals with the memory module of the pontocerebellum. There, large parts of my monograph "Brain Theory of Vertebrates" are taken over almost verbatim. I refrain from identifying them as my own citations, as I hold the copyrights to these texts.
Monograph of Dr. rer. nat. Andreas Heinrich Malczan