Gehirntheorie des Menschen
für das Human Brain Projekt der Europäischen Union
ISBN 978-3-00-068559-0
1. Die Entstehungsgeschichte des Gehirns - das Konzept der Segmente und Neuronenklassen
Das Wirbeltiergehirn entwickelte sich wahrscheinlich aus einem zentralisierten Nervensystem einfachster Mehrzeller. Bei diesen war die Zelldifferenzierung bereits weit fortgeschritten, es gab schon viele Zellarten mit unterschiedlichsten Funktionen.
Im Verlauf der Evolution nahm die Anzahl der Körperzellen stetig zu, so dass die Anzahl der Zellen um ein Vielfaches größer war als die Anzahl der verschiedenen Zellarten. Hier erwies es sich als zweckmäßig, Zellen mit gleicher Aufgabe räumlich zusammenzufassen, so dass sich eine Organbildung manifestierte.
Auch Nervenzellen hatten sich spezialisiert. Die eine Gruppe versorgte die bestehenden Lebenserhaltungssysteme mit Input. Im gesamten Körper wurden über Sensoren wichtige Körpergrößen gemessen und in Feuerraten übersetzt. So wurde beispielsweise die Sauerstoffsättigung überwacht und daraus Steuersignale für die Pumpfrequenz des Herzens abgeleitet. Viele andere Größen, die für die Aufrechterhaltung der Vitalität erforderlich waren, wurden ebenso überwacht.
Die zugehörigen Rezeptoren sandten ihre Signale in die Steuerzentren, die wir als Mittelwertzentren interpretieren. In ihnen wurden die Signale aus dem gesamten Körper zusammengefasst, also eine Art Mittelwert gebildet, der seinerseits das Steuersignal darstellte und entsprechende Reaktionen auslöste. In diesen Mittelwertzentren gab es Mittelwertneuronen, die die ankommenden Signale aus den verschiedenen Teilen des Körpers zusammenfassten und integrierten. Solche Mittelwertzentren finden wir noch heute im menschlichen Gehirn, beispielsweise in der Formatio reticularis.
Wir unterstellen, dass die ersten einfachen Mehrzeller, deren Linien unter anderem zu den Wirbeltieren führten, bereits bilateral symmetrisch waren. Daher war jedes der verschiedenen Mittelwertzentren auf beiden Körperhälften vorhanden. Die Bilateralität blieb auch bei den Wirbeltieren erhalten.
Diese einfachen Mehrzeller waren (unter anderem) die Vorläufer der künftigen Wirbeltiere. Tiere konnten sich – im Gegensatz zu Pflanzen – aktiv bewegen. Wir unterstellen daher die Existenz von Muskeln, die eine Bewegung ermöglichten. Die Muskeln ihrerseits benötigten Steuersignale für ihre Aktivierung. Die Steuersignale wurden von Rezeptoren bereitgestellt. Die Neuronen für den Empfang der sensorischen Steuersignale bildeten auf jeder Körperseite einen eigenen Kern, der als neuronales Organ interpretiert werden kann und den wir als sensorisches Zentrum bezeichnen. Seine Neuronen nahmen den Input der Rezeptoren entgegen, die motorische Aktionen bewirken konnten. Dies waren beispielsweise Tastsignale, olfaktorische Signale, visuelle Signale, aber auch Signale von Rezeptoren, die innerhalb von Muskeln deren Kontraktion maßen.
Der Output des sensorischen Zentrums einer Körperhälfte wurde den Motoneuronen zugestellt, die die Muskeln der gleichen Seite mit Kontraktionssignalen versorgten. Auch diese Neuronen begannen sich räumlich zu konzentrieren und bildeten einen eigenen Neuronenkern, den wir als motorisches Zentrum bezeichnen.
Innerhalb der sensorischen und der motorischen Zentren entstanden Mittelwertneuronen, die mit einem größeren Dendritenbaum die im Zentrum befindliche neuronale Erregung aufnahmen und in die Mittelwertzentren übertrugen. Diese Neuronen interpretieren wir als Mittelwertneuronen, erkennbar am größeren Dendritenbaum. Es war zweckmäßig, die integrierte sensorische und motorische Erregung an die Mittelwertzentren der gleichen Körperhälfte weiterzuleiten. So konnte die Aktivität der Mittelwertzentren an die sensorische und motorische Aktivität des Tieres angepasst werden.
Damit hatten diese Mehrzeller, die zu den Wirbeltieren führten, auf jeder Körperhälfte mehrere Mittelwertzentren, je ein sensorisches und ein motorisches Zentrum.
Es gab bei ihnen mehrere Klassen von Projektionsneuronen, die wir durchnummerieren. Da wir diese Neuronenklassen später im Cortex des menschlichen Gehirns (und im Gehirn aller Wirbeltiere) wiederfinden, geben wir ihnen die Klassennummern, die der Cortexschicht entsprechen, in der wir diese Projektionsneuronen wiederfinden.
Den Mittelwertneuronen in den sensorischen und motorischen Zentren ordnen wir die Klassennummer 6 zu. Wir finden sie in der sechsten Cortexschicht, wo sie große Pyramidenzellen bilden, den Input der Umgebung aufsaugen und an die Mittelwertstrukturen des Gehirns weiterleiten. Sie sind evolutionär gesehen die älteste Neuronenklasse, daher bilden sie im Cortex die unterste, innere Schicht.
Den Projektionsneuronen der sensorischen Zentren ordnen wir die Klassennummer 3 zu. Wir finden sie in der dritten Cortexschicht des Menschen, wo sie den aufsteigenden sensorischen Input aus den Rumpf- und Kopfrezeptoren entgegennehmen und zur motorischen Seite des Cortex weiterleiten. Ebenso finden wir sie in allen Segmenten (Wirbeltiere sind segmentiert), dort empfangen sie den sensorischen Input, der sie über die sensorischen Ganglien erreicht.
Den Projektionsneuronen der motorischen Zentren ordnen wir die Klassennummer 5 zu. Wir finden sie im menschlichen Cortex auf der motorischen Seite in der fünften Schicht. Sie erhalten den Input von den sensorischen Neuronen der Klasse 3, die ihre Axone zu ihnen senden. Weiterhin finden wir sie in allen Segmenten, dort empfangen sie den Output der sensorischen Neuronen und steuern ihrerseits die Motoneuronen an.
Neben den Projektionsneuronen mag es bereits damals Interneuronen gegeben haben, die Interaktionen zwischen Projektionsneuronen ermöglichten. Sie befanden sich stets innerhalb eines neuronalen Kerns. Die hemmenden Interneuronen konnten eine laterale Hemmung ermöglichen. Jedes Projektionsneuron konnte hemmende Interneuronen seiner unmittelbaren Umgebung aktivieren, die ihrerseits die benachbarten Projektionsneuronen hemmten. So standen benachbarte Projektionsneuronen in neuronaler Konkurrenz zueinander und hemmten sich – unter Zwischenschaltung der hemmenden Interneuronen – gegenseitig. So setzten sich stärkere Signale gegen schwächere durch.
Wir unterstellen in dieser Monografie auch einen Signalaustausch zwischen den zwei Körperhälften. Dieser wurde durch Projektionsneuronen der Klasse 2 realisiert. Wir finden sie im Cortex in der zweiten Schicht, wo sie mit ihren Axonen (oft) zur kontralateralen Cortexhälfte (oder in Nachbargebiete) ziehen und ihre Signale abliefern. Sie sind erregend. Ebenso finden wir sie in den Segmenten des Rückenmarks, wo sie die kontralaterale Seite des Segments mit Input versorgen und so beispielsweise motorische Reflexe ermöglichen.
Über die Projektionsneuronen der Klasse 2 standen die zwei Körperhälften in neuronaler Konkurrenz zueinander. Jeder neuronale Kern projizierte über sie erregend in den gleichen Kern der Gegenseite. So projizierte der sensorische Kern über Neuronen der Klasse 2 erregend in den sensorischen Kern der Gegenseite. Die neuronale Konkurrenz entstand, weil diese Signale an hemmenden Interneuronen endeten, die ihrerseits eine laterale Hemmung bewirkten. Gleiches galt für den motorischen Kern jeder Körperseite und für die verschiedenen Mittelwertkerne. Die linke und die rechte Körperhälfte standen in neuronaler Konkurrenz zueinander.
Das war sinnvoll. Stärkere Signale einer Körperhälfte wurden bevorzugt und unterdrückten schwächere Signale der Gegenseite. So fand eine Auswahl statt.
Da das Wachstum der Axone, die die neuronalen Verbindungen realisierten, durch Markersubstanzen gesteuert wurde, musste jede Körperhälfte ihren eigenen Marker besitzen, damit die Projektionsneuronen einer Körperhälfte die Zielstrukturen in dieser Körperhälfte fanden. Daher konnte ein Projektionsaxon der Klasse 2 nicht einfach von einer Körperseite zur kontralateralen Seite ziehen. Es konnte nur zur Trennebene ziehen, welche die beide Körperseiten trennte. Dort musste ein weiteres Projektionsneuron der Klasse 2 die übertragene Erregung entgegennehmen. Weil dieses Neuron sich jedoch bereits auf der Gegenseite befand, wenn auch dicht an der Trennebene, konnte sein Axon die Zielstruktur finden.
Die Signalübergabe von einer Körperseite zur anderen zum Zweck der kontralateralen Hemmung erfolgte also immer über zwischengeschaltete Projektionsneuronen, die sich in der Nähe der Trennebene der beiden Körperhälften befanden. Diese Neuronen bildeten ebenfalls je einen Kern, der als Seitenwechselkern bezeichnet wird. Wir finden solche Seitenwechselkerne im menschlichen Gehirn wieder. Der Nucleus olivaris empfängt den Output des Nucleus ruber und übergibt ihn an das kontralateral gelegene Cerebellum, ist also ein typischer Seitenwechselkern. Die Seitenwechselkerne der beiden Körperhälften befanden sich eng benachbart an der Trennebene der beiden Hälften. Sie konnten im Verlauf der späteren Evolution auch miteinander verschmelzen, so dass dort die Signale beider Körperhälften gleichzeitig zur Verfügung standen und miteinander in Wechselwirkungen treten konnten.
Eine weitere Neuronenklasse könnte damals ebenfalls existiert haben. Die Mittelwertkerne überwachten und steuerten die Lebensfunktionen. Trat durch den Stoffwechsel ein Mangel an den Stoffen auf, die als Energielieferanten dienten, so war Nachschub an Nahrung nötig. Günstig war es, wenn der betreffende Mittelwertkern nicht nur den Energiemangel feststellte, sondern gleichzeitig die Sensorik und die Motorik ankurbelte. Eine erregende Rückprojektion des Mittelwertkerns in das sensorische und motorische Zentrum der jeweiligen Körperseite führte zur Vorerregung der Neuronen. Dieses konnte die Ansprechschwelle senken, auf Reize wurde nun früher reagiert. Möglicherweise konnten Mittelwertkerne sogar Aktionspotentiale in den sensorischen oder motorischen Kernen hervorrufen und den Körper direkt aktivieren. So führte Hunger zur Beutesuche.
Die Einwirkung der Mittelwertkerne auf die sensorischen und motorischen Kerne erfolgte über Neuronen der Klasse 1. Diese Neuronen befanden sich auf jeder Körperseite im sensorischen und im motorischen Kern. Sie empfingen die Mittelwerterregung der Mittelwertzentren der gleichen Körperseite und gaben sie an die Projektionsneuronen des sensorischen und motorischen Kerns weiter, so dass diese aktiviert wurden. Wir finden diese Aktivierungsneuronen im menschlichen Cortex in der ersten Schicht, wo sie mit weit verzweigten Axonen die Umgebungsneuronen aktivieren können. Sie könnten im Verlauf der Evolution als letzte Neuronenklasse entstanden sein, daher finden wir sie im Cortex in der obersten, ersten Schicht. Die Entwicklung dieser Aktivierungsneuronen ermöglichte es, innere Systemzustände zur Aktivierung der Sensorik und Motorik zu verwenden, was einen enormen Fortschritt bedeutete.
Somit besaßen die einfachen Mehrzeller auf einer hinreichend hohen Entwicklungsstufe bereits vier der sechs Neuronenklassen des menschlichen Gehirns. Vorhanden waren damals die Neuronenklassen 1, 2, 3 und 6. Wie kam es zur Entstehung der übrigen zwei Neuronenklassen?
Ich glaube, dass dies beim Übergang vom Mehrzeller zum segmentierten Tier erfolgte.
Wir gehen davon aus, dass es bei den Bilateria, deren Linien zu den Wirbeltieren führten, bei der Vermehrung einen Generationswechsel gab. Die sexuelle Fortpflanzung wechselte sich ständig mit der asexuellen ab. Bei der asexuellen Fortpflanzung stellen wir uns vor, dass sich am Ende des Tieres eine Art Knospe bildete, aus der ein Nachkomme entstand, der sich später vom Muttertier abtrennte. Wenn die Abtrennung unterblieb, entstand ein Doppelwesen, die kleinste Kolonie aus zwei identischen Wesen.
In meiner Monografie „Gehirntheorie der Wirbeltiere“ beschreibe ich dies in Kapitel 2.5 folgendermaßen:
„Zur asexuellen Vermehrung dieser Lebewesen gehörte also die Ausbildung einer Knospe aus der Knospungszone, die sich letztlich zu einem kompletten Zweitlebewesen entwickelte. Der letzte Schritt war die Abtrennung dieses Zweitlebewesens, so dass es ein Nachkomme wurde.
Diese Abtrennung war ein wichtiger Schritt, der in der Erbsubstanz verankert war und biologisch irgendwie gesteuert werden musste. Erfolgte die Codierung der Einzelschritte in der DNA linear in der Reihenfolge ihres Auftretens, so befand sich der Code für die Abtrennung wiederum am Ende der Codierungskette. Möglicherweise ging manchmal dieser Teil bei der Replikation der DNA verloren oder wurde durch besondere Ursachen übersprungen. Dann kam es hier zu einer Störung, und so entstand anstelle eines Nachkommens ein Doppelwesen. War das nicht abgetrennte Tochterwesen danach auch lebensfähig, weil es Nahrung aufnehmen konnte (oder mitversorgt wurde) und über alle nötigen Lebensfunktionen verfügte, so war dieser Reproduktionsfehler nicht sonderlich dramatisch. Es entstand praktisch eine (sehr kleine) Kolonie aus zwei identischen Lebewesen, die miteinander verbunden blieben. Wenn im Verlaufe der Evolution solche Abschnürungsstörungen wiederholt auftraten und sich häuften, konnten sie sich erblich manifestieren. Dann wurde der letzte Schritt des asexuellen Vermehrungsprozesses, die Abschnürung des aus der Knospe entstehenden Nachfolgers, genetisch aus dem Vererbungsprogramm gestrichen und die Entstehung von Zweisegmentern manifestiert.
Man könnte auch davon sprechen, dass hier ein aus zwei Segmenten bestehendes, neues Lebewesen entstanden ist.“
Es ist nicht schwer, sich vorzustellen, wie auf gleiche Art Kolonien aus drei, vier und mehr Tieren entstanden. Wurde diese Koloniebildung ins Vererbungsprogramm aufgenommen und beibehalten, so konnte man ab einer gewissen Evolutionsstufe diese Tiere als segmentierte Tiere auffassen. Sie bestanden zwar aus Einzeltieren, aber spätestens mit der Herausbildung einer Arbeitsteilung zwischen den Segmenten entstanden echte segmentierte Tiere. Bei diesen war die unterbliebene Abtrennung der durch Knospung entstandenen Tochterwesen genetisch manifestiert. Auch im sexuellen Vermehrungsweg entstand ein segmentiertes Wesen. So bildeten sich die segmentierten Bilateria heraus.
Zwischen den Segmenten eines segmentierten Tieres erfolgte ein Stoff- und Signalaustausch. Es entstanden einerseits gemeinsam genutzte Organe bzw. Organsysteme. So etwa der gemeinsam genutzte Darm, der sich anfangs von ersten bis zum letzten Segment erstreckte. Dennoch waren anfänglich die meisten Segmente fast völlig identisch. Einfache Tiere, etwa Würmer, weisen noch diesen ursprünglichen Zustand auf. Doch im Verlauf der Evolution kam es zu starken Veränderungen im Körperbauplan. Welche Gründe mag es dafür gegeben haben?
Ich meine, der Hauptgrund für diese Entwicklung war das ursprüngliche Vorhandensein eines elementaren, zentralen Nervensystems bereits in den Mehrzellern, die später durch Koloniebildung zu segmentierten Tieren wurden. Nur diejenigen Mehrzeller, deren Nervensystem bereits über hemmende Interneuronen zur lateralen Hemmung und zur kontralateralen Hemmung verfügten, konnten einen Entwicklungsweg hin zu den Chordaten einschlagen. Ein weiterer Hauptgrund besteht in der Herausbildung eines neuronalen Signalaustausches zwischen den Segmenten der segmentierten Tiere, aus denen dann die Chordaten hervorgingen.
Dies muss erklärt werden.
Ich habe bereits beschrieben, dass es in den bilateralen Mehrzellern, deren Linien zu den Wirbeltieren führten, die Neuronenklassen 1, 2, 3 und 6 gab, die wir auch in den Cortexschichten des Menschen wiederfinden. Als neuronale Strukturen besaßen diese Tiere auf jeder Körperseite ein sensorisches Zentrum, ein motorisches Zentrum und mehrere, verschiedene Mittelwertzentren. Letztere dienten den Lebenserhaltungssystemen als Inputlieferanten, wirkten jedoch auch aktivierend auf die sensorischen und motorischen Zentren zurück.
Mit der Entstehung von segmentierten Bilateria fand man den beschriebenen Aufbau in jedem Segment wieder, da (fast) alle Segmente anfänglich identisch waren. Nur das letzte Segment verfügte bei diesen Tieren zusätzlich über eine Knospungszone zur asexuellen Vermehrung.
Ich gehe davon aus, dass die neuronalen Signale der Rezeptoren eines Segments im Verlauf der Evolution die Segmentgrenzen überwinden konnten. Dies lag an der engen Verwandtschaft benachbarter Segmente, die sich auf die Neuronen übertrug. Noch heute beobachtet man, dass die Rezeptorsignale segmentierter Lebewesen die Segmentgrenzen überschreiten können und in Nachbarsegmenten enden. Ein Beispiel hierfür sind die Dermatome des Menschen. Ein Dermatom stellt den Hautbereich dar, der von den sensiblen Fasern einer Spinalnervenwurzel autonom versorgt wird. Die Dermatome überwinden ebenfalls die Segmentgrenzen, so dass sie sich überlappen. Hier erreicht die Information aus einem Segment auch die Nachbarsegmente.
Für den Signalaustausch zwischen den Segmenten entwickelten sich im Verlauf der Evolution spezielle Neuronenklassen. Eine Neuronenklasse übernahm den Signaltransport in Richtung des Kopfes, also die afferente Richtung. Wir ordnen dieser Neuronenklasse die Klassennummer 4 zu. Wir finden sie im menschlichen Cortex in der vierten Schicht. Dort sind ihre Neuronen jedoch zu Interneuronen geworden, weil der Cortex dem höchsten kopfseitigen Segment entspricht und somit kein höheres Segment existiert, an das diese Neuronen der Klasse 4 ihre Erregung weitergeben könnten.
Den Signalaustausch in Richtung des Schwanzes übernehmen Projektionsneuronen, denen wir die Neuronenklasse 5 zuordnen. Wir finden sie in der fünften Cortexschicht des Menschen. Dort übernehmen sie die Cortexerregung und projizieren schwanzwärts zu den Motoneuronen.
Damit wird eine Aufgabenteilung zwischen den Neuronen der Klasse 4 und 5 deutlich. Die Neuronen der Klasse 4 sind sensorische Projektionsneuronen. Sie übertragen die Signale aus dem sensorischen Zentrum in die sensorischen Zentren der übergeordneten Segmente.
Die Neuronen der Klasse 5 sind motorische Neuronen. Sie übertragen die Signale aus den motorischen Zentren schwanzwärts zu den Motoneuronen derjenigen Segmente, denen die jeweiligen motorischen Signale zugeordnet sind.
Ein Grund für die Aufspaltung der vertikalen Signalübertragung in zwei voneinander unabhängige Arten lag in der Steuerung der Wachstumsrichtung der Axone durch Marker. Die Axone der Klasse 4 hatten die entgegengesetzte Wuchsrichtung der Axone der Klasse 5. Vermutlich reagierten sie auf einen Kopfmarker, während die anderen dem Schwanzmarker folgten.
Und weil der Körper dieser Tiere bilateral symmetrisch ist, bilden die Axone der sensorischen Neuronen der Klasse 4 auf jeder Körperhälfte einen Nervenstrang, der vom Schwanzsegment zum Kopfsegment verlauft und in jedem Segment je ein Ganglion bildet, welches das sensorische Zentrum des Segments enthält.
Ebenso bilden die Axone der motorischen Neuronen der Klasse 5 auf jeder Körperseite einen Nervenstrang, der vom Kopfsegment zum Schwanzsegment verläuft und in jedem Segment ein Ganglion bildet, welches das motorische Zentrum des Segments enthält.
Da in jedem Segment – wie beim zuvor beschriebenen Mehrzeller – das sensorische Zentrum mit dem motorischen durch die Axone der Neuronen der Klasse 3 verknüpft ist, sieht das Nervensystem dieser Bilateria auf jeder Körperseite so aus wie eine Strickleiter. Daher wird es auch im deutschen Sprachraum als Strickleiternervensystem bezeichnet.
Beide Strickleitern der zwei Körperhälften sind zusätzlich durch die Axone der Neuronen der Klasse 2 verknüpft, die jeweils das linke und rechte motorische bzw. das sensorische Zentrum verbinden und an hemmenden Interneuronen enden, um die kontralaterale Hemmung zu realisieren. Bei Übergang von einer Körperseite zur anderen wird pro Segment und Körperseite ein Seitenwechselkern benötigt. Insgesamt sind vier Nervenstränge beteiligt, zwei sensorische und zwei motorische. Somit liegt hier das klassische tetraneurale Nervensystem vor.

Erwähnen sollten wir hier, dass es in jedem Segment die Mittelwertsysteme gibt, die sensorischen und motorischen Input empfangen und aktivierend in Neuronen der Klasse 1 projizieren, die sowohl im motorischen als auch im sensorischen Zentrum vorhanden sind. Auch hier entwickelte sich ein Signalaustausch zwischen den Segmenten. Wir gehen vereinfachend davon aus, dass hierfür auch die Neuronen der Klassen 4 und 5 verwendet wurden.
Im Verlauf der Evolution wuchs die Körpergröße der Tiere, die Anzahl der Rezeptoren wuchs ebenso und es entwickelten sich auch viele neue Rezeptorenarten. Dies führte zu einer starken Zunahme der Anzahl der signalverarbeitenden Neuronen in den sensorischen und motorischen Zentren sowie in den Mittelwertzentren. Damit verbunden war ebenso ein starker Anstieg der Anzahl derjenigen Neuronen, die den Signalaustausch zwischen den Segmenten ermöglichten.
Die Anzahl der Axone der Projektionsneuronen der Klassen 4 und 5 wuchs so stark, dass zunächst breite Bänder aus Axonen den kopf- und schwanzseitigen Signaltransport bewerkstelligten. Diese verbreiterten sich, und nahmen eine Form an, die ebenso wurmähnlich war wie der Körper der damaligen Tiere. So entstand das sogenannte Neuralrohr. Es besteht zur Hälfte aus den Axonen der kopfwärts projizierenden Neuronen der Klasse 4, während die andere Hälfte aus den Axonen der schwanzwärts projizierenden Neuronen der Klasse 5 besteht. Während sich die Axone jeweils außen anordnen, nehmen die zugehörigen Zellkörper der Neuronen den inneren Platz ein und bilden dort die graue Substanz. Als Neuralrohr wird dieses System deshalb bezeichnet, weil sich im Inneren ein flüssigkeitsgefüllter Ventrikelraum befindet. Die Zellkerne suchen die Nähe dieses Ventrikelraumes, möglicherweise weil sie ihre Nährstoffe von ihm beziehen und ihre Stoffwechselprodukte an ihn abgeben.
Im Neuralrohr, aber auch im späteren Rückenmark enden die sensorischen Rezeptoren des Körpers der Tiere in der sensorischen Hälfte, die als Hinterhorn bezeichnet wird, während die Motoneuronen in der als Vorderhorn bezeichneten Neuralrohrhälfte angeordnet sind. Die sensorischen Neuronen der Klasse 3 projizieren über ihre Axone in die motorische Hälfte, dabei wird die jeweilige Körperseite beibehalten. Dies ist in der folgenden Abbildung dargestellt, die Neuronen der Klasse 2 zur kontralateralen Hemmung sind vereinfachend weggelassen worden.
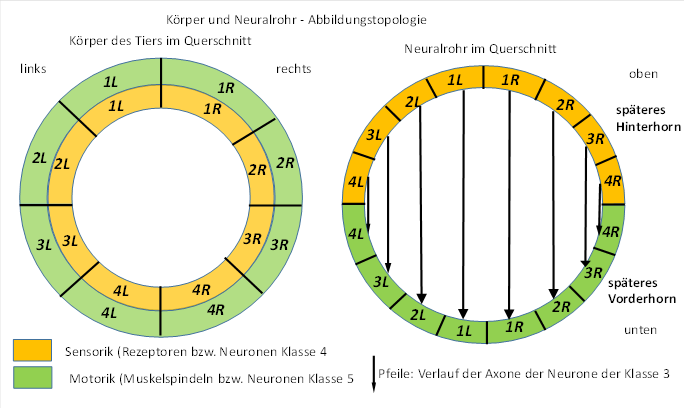
Abbildung 2: Körper und Neuralrohr - Abbildungstopologie
Bildquelle: Gehirntheorie der Wirbeltiere, Andreas Malczan, 2020, Abb. 6
Der Ventrikelraum führt zur Krümmung der Axone der Neuronenklassen 2 und 3, da diese nicht in ihn eindringen können (In obiger Abbildung nicht berücksichtigt).
Im Neuralrohr derjenigen Lebewesen, deren Linie zu den Wirbeltieren führte, entstand nach meiner Ansicht eine Wohlordnung der verschiedenen Neuronenklassen in eigenständigen Schichten von außen nach innen in folgender Reihenfolge:
· Schicht 6 der Neuronenklasse 6 (Mittelwertneuronen)
· Schicht 5 der motorischen Kommissurneuronen der Klasse 5
Schicht 4 der sensorischen Kommissurneuronen der Klasse 4
Schicht 3 der sensorischen Konnektivneuronen der Klasse 3
Schicht 2 der motorischen Konnektivneuronen der Klasse 2
Schicht 1 der Aktivierungsneuronen der Klasse 1
Hierbei befindet sich die Schicht 1 mit den Neuronen der Klasse 1 unmittelbar benachbart zum Ventrikelraum des Neuralrohrs. Die Nachbarschaft des Ventrikelraumes zur Neuronenklasse 1 scheint für die Schichtungsrichtung wesentlich zu sein.
Es ist auffällig, dass die Neuronen im menschlichen Cortex genau in der umgekehrten Reihenfolge in Schichten angeordnet sind. Dies liegt sicherlich an der räumlichen Kopplung der Neuronen der Klasse 1 an das äußere Ventrikelsystem des Cortex.
In jedem neuronalen Kern existieren nach unserer Übereinkunft hemmende Interneuronen zur lateralen Hemmung. Genau diese Tatsache ist ein extrem wichtiger Grund für die Spezialisierung der Segmente. Findet man anfänglich noch fast jedes Organ in fast jedem Segment, so trifft dies etwa bei Wirbeltieren nicht mehr zu. Wie ist dies zu erklären?
Dies hängt mit der Organisationsform der Signalweiterleitung zusammen. Wir unterteilen den Input eines Segments in Eigeninput und Fremdinput. Der Eigeninput entstammt denjenigen Rezeptoren, die in diesem Segment vorhanden sind. Fremdinput liefern die Rezeptoren aller übrigen Segmente, die schwanzwärts angeordnet sind.
Wir unterstellen hier, dass der Fremdinput in jedem Segment so behandelt wurde wie der Eigeninput. Der Eigeninput endete an Neuronen der Klasse 3, die zum motorischen Zentrum projizierten, aber ebenso an Neuronen der Klasse 4, die zum sensorischen Zentrum des benachbarten Segments in Kopfrichtung zogen. Der Fremdinput, der das sensorische Zentrum über aufsteigende Axone der Projektionsneuronen der Klasse 4 erreichte, wurde ebenso behandelt. Er endete einerseits an Neuronen der Klasse 3, die zum motorischen Zentrum des gleichen Segments auf der gleichen Körperseite projizierten. Andererseits endete er an sensorischen Projektionsneuronen der Klasse 4, die zum kopfseitigen Nachbarsegment zogen und dort im sensorischen Zentrum endeten.
Damit empfing bei denjenigen segmentierten Bilateria, deren Linien zu den Wirbeltieren führten, jedes Segment eine Kopie aller sensorischen Signale derjenigen Segmente, die in Schwanzrichtung vorhanden waren. Das erste Kopfsegment erhielt den sensorischen Input aus allen Segmenten des Tieres. Dort war die meiste Information vorhanden, dort bildete sich der Cortex heraus. Dies wird später ausführlicher beschrieben werden.
Was passierte jedoch mit den motorischen Signalen?
Der Eigeninput eines Segments kam im sensorischen Zentrum an und wurde zum motorischen Zentrum durchgereicht. Dort traf er auf die Neuronen der Klasse 5. Diese projizierten in die Motoneuronen dieses Segments. Damit hatten die Eigensignale ihre Ziele erreicht.
Der Fremdinput entstammte anderen, tiefer gelegenen Segmenten. Die motorischen Ziele befanden sich jeweils im Herkunftssegment der Signale. Daher wurde der Fremdinput über abwärts projizierende Neuronen der Klasse 5 zum Nachbarsegment weitergereicht. Dort wurde der Teil der Signale, die ursprünglich von den Rezeptoren dieses Segments geliefert worden waren, an die zugeordneten Projektionsneuronen der Klasse 5 übergeben, die ihn an die Motoneuronen des Segments weitergaben. Die übrigen Signale des Fremdinputs wurden absteigend über Neuronen der Klasse 5 von Segment zu Segment übergeben, bis sie ihr Herkunftssegment erreicht hatten und dort an die zugeordneten Motoneuronen weitergegeben werden konnten.
Im Kopfsegment lag der motorische Output noch für alle Segmente vor. Von Segment zu Segment wurde dieser komplette Output vermindert jeweils um die motorischen Signale, die im aktuellen Segment ihre motorischen Ziele besaßen. Das Schwanzsegment erhielt nur noch Signale, die seinen eigenen Motoneuronen zugeordnet waren.
Diese besondere Organisationsform sollte sich für die Wirbeltiere als besonders nützlich erweisen. Sie konnte jedoch in anderen Tieren in ähnlicher Form ausgebildet werden.
Wenn die neuronalen Signale während ihrer Ausbreitung auf den Axonen der sensorischen oder motorischen Projektionsneuronen gedämpft wurden, also mit zunehmender Wegstrecke schwächer wurden, erreichten sie nicht mehr alle Segmente. Dann reichte der Signalaustausch nur über wenige Nachbarsegmente. Bei Würmern ist ein solcher Zustand zu beobachten. Dies verhinderte eine weitere evolutionäre Entwicklung in Richtung Wirbeltiere.
Der sensorische Signalaustausch definierte eine Nachbarschaftsbeziehung, in der es untergeordnete und übergeordnete Segmente gab. Alle schwanzseitig gelegenen Segmente waren untergeordnet, alle kopfseitig gelegenen übergeordnet.
Jedes Segment besaß neben seinem eigenen sensorischen Input auch den Input aller untergeordneten Segmente.
Hier ist es an der Zeit, sich diese neuronale Organisationsform genauer zu betrachten. Wie waren die Neuronen räumlich angeordnet?
Im Prinzip ist es am einfachsten, wenn wir uns vorstellen, jedes Segment besäße neben seinem eigenen sensorischen Zentrum eine Kopie aller sensorischen Zentren der untergeordneten Segmente. Eine Ansammlung von sensorischen Zentren. Jede Kopie wäre so aufgebaut wie ihr Original. Im Kopfsegment befänden sich dann so viele sensorische Zentren pro Körperhälfte, wie es Segmente gab.
Damit vollziehen wir den Übergang vom sensorischen Zentrum zum Ganglion. Ein sensorisches Ganglion auf einer Körperseite eines Segments besteht aus dem sensorischen Zentrum dieses Segments und aus den Kopien der sensorischen Zentren aller untergeordneten Segmente. Und das Strickleitersystem besteht aus solchen Ganglien.
Auf der motorischen Seite war es ebenso. Im Kopfsegment gab es das motorische Zentrum des Kopfsegments sowie die Kopien aller motorischen Zentren der untergeordneten Segmente. Sie bildeten das motorische Ganglion. Und in jedem tiefer gelegenen Segment gab es das motorische Zentrum dieses Segments sowie die Kopie der motorischen Zentren aller untergeordneten Segmente.
Diese recht einfache Organisationsform war sicherlich einfach in eine Befehlsfolge umzusetzen, die in der DNA des Tieres die Bauanleitung für das Nervensystem lieferte.
Nun müssen wir nur noch klären, wie die Kopien der neuronalen Zentren in jedem Segment angeordnet waren. Hierzu stellen wir uns bereits einen höheren Zustand der Entwicklung vor und fordern eine möglichst einfache Organisationsform. Betrachten wir beispielhaft die rechte Körperseite des Tieres. Dann möge sich das sensorische Zentrum rechts außen befinden, während links von ihm die Kopie des sensorischen Zentrums des schwanzseitigen Nachbarsegments angelagert wird. Links daneben befindet sich die Kopie des sensorischen Segments des nächsten, schwanzseitigen Segments. Das wiederholt sich solange, bis die Kopie des Schwanzsegments angelagert wird.
Innen befinden sich die Kopien der schwanzseitigen Segmente, wohlgeordnet nach aufsteigenden Segmenten, außen liegt das sensorische Zentrum des Segments.
Bei den motorischen Zentren gibt es eine entsprechende Wohlordnung. Jedes motorische Ganglion besteht aus der Aneinanderreihung von motorischen Zentren, von denen nur das dem Segment zugehörige als Original vorliegt, die übrigen stellen Kopien der motorischen Zentren der untergeordneten Segmente dar. Benachbarte Segmente projizieren in benachbarte Kopien.
Eine weitere Sortierung fand statt, als sich verschiedene Arten von Rezeptoren bildeten. Wir können davon ausgehen, dass eine neue Rezeptorenart sich meist aus einer bestehenden Rezeptorenart bildete. Dann spaltete sich die zugehörige Modalität in zwei neue Modalitäten. Wir unterstellen, dass sich die Projektionsneuronen dann ebenfalls räumlich trennten.
Mit der Zunahme der Rezeptorenarten war ebenso eine Zunahme der Anzahl der Rezeptoren verbunden. Rezeptoren einer Rezeptorenart, etwa Tastrezeptoren, waren ab einer gewissen Evolutionsstufe überall in der Körperoberfläche verteilt.
Die zunehmende Anzahl der Rezeptoren und der Rezeptorenarten führten zum Übergang vom strickleiterartigen Nervensystem zu einem röhrenförmigen. Grund war der röhrenförmige, wurmähnliche Körperbau dieser frühen Tiere. So konnte im Neuralrohr die Körpertopologie einfach nachgebaut werden. Die Axone der sensorischen Projektionsneuronen bildeten zusammen mit den Axonen der motorischen Projektionsneuronen ein Neuralrohr, dessen Inneres von einem flüssigkeitsgefüllten Ventrikelraum gefüllt wurde. Die als Hinterhorn bezeichnete Hälfte bestand aus Neuronen der Klasse 1, 2, 3, 4 und 6 sowie aus ihren Axonen. Die Axone der sensorischen Projektionsneuronen der Klasse 4 sind im Neuralrohr des Menschen mit Myelin umgeben. Dieses erhöht die Reichweite der neuronalen Signale. Die Axone verlaufen im Neuralrohr stets außen, dort ist die weiße Substanz angeordnet. Die weiße Farbe wird vom Myelin hervorgerufen.
Innen im Hinterhorn befinden sich einerseits die Zellkerne der Projektionsneuronen der Klassen 1, 2, 3 und 4 sowie die nicht myelinisierten Axone der Neuronen der Klasse 3, die zu den motorischen Neuronen projizieren.
Im Vorderhorn des Neuralrohrs befinden sich die Zellkerne der motorischen Projektionsneuronen der Klasse 1, 2, 5 und 6 und deren Axone, die zu den Motoneuronen in den verschiedenen Segmenten projizieren. Zwischen Vorderhorn und Hinterhorn verlaufen die Querverbindungen, die von den Neuronen der Klassen 3 und 2 gebildet werden.
Neben Vorderhorn und Hinterhorn gibt es im oberen Bereich noch das Seitenhorn, welches zwischen beiden liegt. Es wird von den Zellkörpern und Axonen der vegetativen Neuronen gebildet wird und gehört zum Sympathikus.
Auch im Neuralrohr beobachtet man eine topologische Wohlordnung. Es gibt eine Ordnung nach Segmenten und nach Modalitäten.
Die Projektionsaxone der Neuronen der Klasse 4 der unteren Segmente verlaufen innen. Höhere, kopfseitige Segmente lagern ihre Axone seitlich außen an.
Im Hinterhorn ist es analog. Axone der Neuronen der Klasse 5, die zu schwanzseitigen Segmenten projizieren, befinden sich innen, auch hier lagern höhere, kopfseitige Segmente ihre Axone außen an.
So ergibt sich bezüglich der afferenten und efferenten Projektionen eine Art Baumringstruktur. Axone der unteren Segmente bilden die inneren Ringe, während Axone der höheren Segmente äußere Ringe bilden.
Doch auch hinsichtlich der signalliefernden Rezeptoren bzw. der signalempfangenden Motoneuronen gibt es eine topologische Wohlordnung. Die Rezeptoren, die den sensorischen Teil, also das Hinterhorn, mit Signalen versorgen, entstanden im Verlauf der Evolution schrittweise nacheinander. Die Axone, die Signale der evolutionär ältesten Rezeptoren kopfwärts transportieren, sind innen angeordnet, die der evolutionär jüngeren dagegen außen. So bilden die verschiedenen Modalitäten ringförmige Strukturen, die wir als Modalitätenringe bezeichnen. Ein Modalitätenring besteht aus einem sensorischen Halbring, gebildet von den afferent projizierenden Axonen der Klasse-4-Neuronen und einem motorischen Halbring. Die Neuronen, die ihn bilden, empfangen ihre Signale über die Querverbindungen der Klasse 3, die von den Neuronen der Klasse 4 mit Input beliefert werden.
Zusätzlich sind die Modalitätenringe in Segmentringe unterteilt. Innere Segmentringe gehören zu tieferen, schwanzseitig liegenden Segmenten.
Diesen Grundaufbau des Neuralrohrs wollen wir als Startmodell wählen, um die künftige Entwicklung des zentralen Nervensystems der Wirbeltiere, Säugetiere, Primaten und letztlich des Homo sapiens zu analysieren.
Gewiss gab es Bilateria, die diesen neuronalen Aufbau nie erreichten. So war es möglich, dass bei vielen Arten die neuronale Reichweite der Signale nur wenige Segmente überwinden konnte. Dann erhielt das oberste Segment nicht die komplette Information des gesamten Körpers aus allen Segmenten, sondern nur den Input aus einem oder zwei Nachbarsegmenten. Diese Tiere konnten keine Wirbeltiere werden, bei denen das oberste Segment über den kompletten Input aus allen Segmenten verfügte.
Doch die Tiere, deren Neuralrohr so wie beschrieben aufgebaut ist, waren den Wirbeltieren noch sehr unähnlich. Insbesondere waren alle Segmente noch ziemlich identisch aufgebaut. In fast allen Segmenten waren noch fast alle Organe vorhanden. Um sich letztlich zum Wirbeltier zu entwickeln, musste nicht nur eine Wirbelsäule entstehen. Viele Organe, die zuvor in jedem Segment vorhanden waren, musste sich zurückbilden, so dass sie letztlich nur noch in einem oder wenigstens in wenigen Segmenten übrigblieben. Wir werden diesen Prozess als Reduktion bezeichnen. Was könnte diese Entwicklung begünstigt haben?
Nach meiner Ansicht verdanken wir diese Entwicklung der lateralen Hemmung. In jedem Segment und in jedem seiner neuronalen Kerne gab es hemmende Interneuronen, die der Kontrastverstärkung dienten. Diese wurde durch die gegenseitige Hemmung der beteiligten Neuronen mittels der hemmenden Interneuronen erreicht. Hemmende Interneuronen ermöglichten neuronale Konkurrenz. Und bei Konkurrenz gibt es Gewinner und Verlierer.
Betrachten wir beispielhaft die Rezeptoren des Riechsinns, der auch als olfaktorischer Sinn bezeichnet wird. Er ist einer der ältesten Sinne. Bereits die Mehrzeller, aus denen sich später durch Koloniebildung die segmentierten Bilateria bildeten, besaßen olfaktorische Rezeptoren. Kein Wunder, dass diese bei den frühen segmentierten Tieren in (fast) allen Segmenten vorhanden waren, denn anfangs waren die Segmente ziemlich identisch. Jeder olfaktorische Rezeptor lieferte sein Signal an ein angeschlossenes Neuron der Klasse 3 sowie an ein Neuron der Klasse 4 ab. Das Neuron der Klasse 3 projizierte in ein motorisches Neuron der Klasse 5, welches ein Motoneuron in diesem Segment kontaktierte. Das Neuron der Klasse 4 projizierte dagegen kopfwärts. Im kopfseitigen Nachbarsegment kontaktierte es wiederum ein Neuron der Klasse 4, welches zum nächsten kopfseitigen Nachbarsegment projizierte. So kam dieses Signal letztlich im Kopfsegment an. Dort befand sich bereits eine Unmenge von sensorischen Neuronen, welche die Signale genau dieser Modalität aus den verschiedensten Segmenten empfingen. Denn jedes Segment sendete die olfaktorischen Rezeptorsignale in alle ihm übergeordneten Segmente, also auch in das Kopfsegment.
Die Neuronen, die die Signale empfingen, aktivierten ihrerseits hemmende Interneuronen, die eine laterale Hemmung verursachten. Damit hemmte jedes olfaktorische Signal die olfaktorischen Signale der benachbarten Neuronen. Alle Segmente des Körpers standen daher bezüglich der Olfaktorik in neuronaler Konkurrenz zueinander. Die stärkste Konkurrenz lag im Kopfsegment vor. Hier trafen ja alle olfaktorischen Signale aus allen Segmenten ein.
Die Signale aus dem Schwanzsegment wurden durch die laterale Hemmung am stärksten betroffen, denn die olfaktorischen Signale aller übrigen Segmente wirkten hemmend auf sie ein. Dies lag auch daran, dass die olfaktorischen Rezeptoren in den verschiedenen Segmenten von einem Duft gleichzeitig erregt wurden. Die Tiere lebten damals im Urozean, ein Duft breitete sich im Wasser schnell nach allen Seiten aus. So waren alle olfaktorischen Rezeptoren aller Segmente, die auf einen vorhandenen Duft reagierten, gleichzeitig aktiv. Das traf besonders dann zu, wenn die Körper der Tiere noch relativ klein waren.
Dies führte über einen längeren evolutionären Zeitraum zur neuronalen Atrophie. Neuronen, die ständig unterdrückt werden, bilden sich zurück und verkümmern. Am Ende des Prozesses sind sie nicht mehr vorhanden. So verschwanden die olfaktorischen Rezeptoren im Schwanzsegment komplett. Im Verlauf der Evolution wurden im letzten Segment gar keine olfaktorischen Rezeptoren angelegt, das genetische Programm hatte sich geändert.
Doch auch die Mittelwertkerne waren von dieser Entwicklung betroffen. Das olfaktorische Mittelwertzentrum, welches nach meiner Ansicht beim Menschen von den Septumkernen (Nuclei septales) gebildet wird, empfing im Schwanzsegment nach der Atrophie der olfaktorischen Rezeptoren auch keinen Input mehr, so dass es ebenfalls atrophierte.
Damit war die Entwicklung nicht abgeschlossen, denn nun konkurrierten in den verbliebenen Segmenten – das Schwanzsegment ausgenommen – wieder alle olfaktorischen Signale um die Vorherrschaft. Neuronale Konkurrenz kennt Sieger und Verlierer. Der Gewinner war das Kopfsegment, die Verlierer waren die übrigen Segmente. Im Verlauf eines länger andauernden Evolutionsprozesses atrophierten alle olfaktorischen Rezeptoren in allen nachfolgenden Segmenten. Damit hatte sich das Kopfsegment auf die Olfaktorik spezialisiert, die übrigen Segmente besaßen weder olfaktorische Rezeptoren, noch verfügten sie über septale Kerne, die als olfaktorische Mittelwertkerne fungierten.
Aber die neuronale Konkurrenz war noch viel tiefgreifender. Anfänglich waren die Neuronen noch nicht so spezifiziert wie in späteren Evolutionsepochen. Sie unterschieden sich noch nicht wesentlich. Es war – vereinfacht gesprochen – den Neuronen völlig egal, durch welche Rezeptoren sie erregt wurden. Eine besondere Spezifität hinsichtlich der Modalität bestand anfangs nicht oder nur in geringem Maß.
Daher konkurrierten im Kopfsegment alle Signale untereinander, also beispielsweise olfaktorische mit visuellen, motorischen und anderen. Jedes Signal stand mit jedem Signal der gleichen Körperhälfte in Konkurrenz. Da aber die olfaktorischen Signale bereits (vielleicht durch Zufall – oder weil sie die Ersten waren) die Oberhand gewonnen hatten, gerieten die übrigen Modalitäten unter Druck. Die laterale Hemmung der Signale untereinander und die größere Durchsetzungskraft der olfaktorischen Signale führte dazu, dass die Rezeptoren der übrigen Modalitäten im Kopfsegment zurückgebildet wurden, atrophierten und letztlich völlig verschwanden. Die Olfaktorik blieb im Kopfsegment der Sieger und blieb als alleinige Modalität im Kopfsegment übrig. Dass gerade die Olfaktorik den Wettbewerb auf Grund der lateralen Hemmung gewann, mag auch daran gelegen haben, dass sie einer der frühesten Sinne war, die die Tierwelt hervorgebracht hatte. Die Mittelwertkerne in Kopfsegment konnten nur noch olfaktorische Signale empfangen. Mittelwertkerne, die von anderen Signalarten versorgt wurden, verschwanden aus dem Kopfsegment.
Es ist nicht schwer, sich vorzustellen, dass im zweiten Segment die Sehrezeptoren den multimodalen Konkurrenzkampf der Segmente und Modalitäten gewannen. So wurde im zweiten Segment der visuelle Sinn zum alleinigen Gewinner, alle übrigen Modalitäten verschwanden hier. Und in allen nachfolgenden Segmenten wären die visuellen Rezeptoren auch durch Atrophie verschwunden, gäbe es da nicht den Vestibularsinn. Er ist der Grund dafür, dass es neben dem Augenpaar im zweiten Segment noch ein weiteres Augenpaar gab.
Doch diejenigen Tiere, die den bisher beschriebenen Stand der Evolution erreicht hatten, müssen schon Augenmuskeln besessen haben. Standardmäßig waren sechs Augenmuskeln in jedem Auge vorhanden.
Bereits Spinnen, die zu den einfacheren segmentierten Tieren zählen, verfügen über Augen. Hier trat noch keine vollständige Reduktion der Augenanzahl durch die laterale Hemmung auf, so dass mehrere Segmente je ein Augenpaar besitzen. Die meisten Spinnen besitzen acht Augen. Man unterscheidet Hauptaugen und Nebenaugen. Bei den Hauptaugen der Spinnen gibt es Augenmuskeln. Während bei Wirbeltieren die Augenmuskeln das Auge von außen bewegen, kann bei Spinnen mit Hilfe der Augenmuskeln die Netzhaut, also die Retina, seitlich verschoben werden. Bei vielen Spinnen sind es pro Hauptauge sechs Augenmuskeln, jedoch weisen etliche Arten auch eine geringere Anzahl auf. Möglicherweise ist dies bereits eine Rückbildung, denn der Sehsinn ist bei vielen Spinnen von untergeordneter Bedeutung.
Der Besitz von sechs (äußeren) Augenmuskeln bei den segmentierten Tieren, deren Linien zu den Wirbeltieren führten, wird hier vorausgesetzt. Es gibt einen triftigen Grund dafür.
Im dritten Segment, welches direkt an das zweite, visuelle Segment grenzte, konnten sich die Augenmuskelsignale durchsetzen und verdrängten alle anderen Rezeptorarten dieses Segments, ebenso die nichtvisuellen Mittelwertzentren. So entstand im dritten Segment im sensorischen Zentrum eine neuronale Struktur, die wir später bei den Wirbeltieren als Tectum opticum, kurz als Tectum bezeichnen. Hier überlagern sich visuelle Signale mit den Signalen der Muskelspannungsrezeptoren aus den sechs (äußeren) Augenmuskeln. Das Tectum entwickelte sich bei Wirbeltieren zu einem bedeutsamen visuellen Zentrum.
Ein siebenter äußerer Augenmuskel, der Musculus retractor bulbi („Zurückzieher des Auges“) fehlt beim Menschen, ist aber bei den meisten übrigen Säugetieren ausgebildet. Er dient nicht der Augenbewegung an sich, sondern dem Schutz des Auges bei Annäherung an Hindernisse.
Durch die sechs äußeren Augenmuskeln ist es möglich, die Augen stets so auszurichten, dass wichtige visuelle Objekte gut erfasst und analysiert werden können. Der Algorithmus wird später beschrieben werden. Als prinzipielles Modul kommt hier das Schwerpunktmodul zur Anwendung. Später ermöglicht ein Differenzmodul sogar das Bewegungssehen. Doch dazu später. Hier erklären wir zunächst die schrittweise Weiterentwicklung der segmentierten Bilateria zu Wirbeltieren.
Nachdem sich im ersten Segment der olfaktorische Sinn durchgesetzt hatte, im zweiten der Sehsinn, im dritten die Rezeptoren der Augenmuskeln, ging die Reduktion durch die laterale Hemmung weiter.
Ursprünglich gab es in jedem Segment den Vestibularsinn. Er wertete mit Hilfe der Schwerkraft die Lage des Lebewesens im Raum aus. Bereits die einfachen Mehrzeller, die noch gar nicht segmentiert waren, besaßen ihn. Beispielsweise Polypen, die ihn nutzten, um ihre Körperachse senkrecht auszurichten.
In einem flüssigkeitsgefüllten Hohlraum, der Statocyste, rollte ein Sandkorn immer an die tiefste Stelle. Die Wandung der Statocyste war mit Haarzellen ausgekleidet. Diese wurden durch das Gewicht des Sandkorns verbogen, so dass sie Aktionspotentiale erzeugten. Diese aktivierten die Muskeln, welche die Stellung des Körpers im Wasser so korrigierten, dass die Standardlage wieder eingenommen wurde. Dazu musste der Schwanz des Körpers entsprechend gebogen werden. Die Muskeln wurden von den Aktionspotentialen der Statocyste aktiviert.
Diese neuronale Schaltung erforderte zwingend eine Signalkreuzung. Es mussten immer die Muskeln der kontralateralen Körperseite aktiviert werden.
Als sich die segmentierten Tiere herausgebildet hatten, besaß anfänglich jedes Segment zwei Statocysten, jeweils eine auf der linken und eine auf der rechten Körperseite. Die neuronale Konkurrenz aller Signale und aller Segmente untereinander führte dazu, dass sich im ersten Segment die olfaktorischen Signale, im zweiten die Signale der Sehrezeptoren und im dritten die Signale der Augenmuskeln durchgesetzt hatten und die übrigen Rezeptoren in diesen drei Segmente verdrängten.
So kam es, dass die Statocysten im ersten, zweiten und dritten Segment den Konkurrenzkampf verloren und letztlich degenerierten. Ab einer gewissen Evolutionsstufe gab es in den ersten drei Segmenten keine Statocysten mehr. Die Statocysten in den übrigen Segmenten traten aber ebenfalls in den neuronalen Konkurrenzkampf. Nun hemmten die Statocysten im vierten Segment die übrigen Statocysten am erfolgreichsten, so dass in den übrigen, schwanzseitig gelegenen Segmenten eine schrittweise Rückbildung der Statocysten begann. Dies erfolgte über viele hundert, tausend oder gar Millionen von Generationen und verlief schrittweise.
Anfänglich wurde die kleine Öffnung, durch die zunächst ein Sandkorn in die Statocyste eindrang, größer. Das Sandkorn fiel heraus. Die Öffnung wurde noch größer, so dass das Wasser von außen in die Statocyste eindringen konnte. Nun jedoch reagierten die Haarzellen nicht mehr auf das Gewicht des Sandkorns – dieses gab es nicht mehr. Stattdessen reagierten sie auf die Wasserströmung, die durch die Bewegung des Tieres verursacht wurde. Dies war der Beginn der Entwicklung des Seitenliniensystems der Wassertiere. Die Wasserströmung, die vom Tier durch seine Eigenbewegungen verursacht wurde, konnte neuronal bewertet werden. Viel wichtiger jedoch war die Wasserströmung, die durch bewegte Fremdobjekte verursacht wurde. Fressfeinde verursachten solche Wasserströmungen bei Annäherung, aber auch Tiere oder Pflanzen, die auf dem eigenen Speiseplan standen. So wurde das Seitenliniensystem ein wichtiger Fernortungssinn. Sehr brauchbar vor allem auch in tiefem Wasser, das vom Licht nicht durchdrungen werden konnte. Das Seitenliniensystem war ab dem fünften Segment in allen nachfolgenden Segmenten vorhanden, während die vestibulären Statocysten nur noch im vierten Segment übrigblieben.
Warum führte die neuronale Konkurrenz der Rezeptoren des Seitenliniensinns nicht zur Atrophie in allen Segmenten, die auf das fünfte Segment folgten?
Eine Atrophie konnte nur stattfinden, wenn Neuronen ständig gleichartige (identische) Signale empfangen. Nur dann sind sie immer gleichzeitig synchron aktiv und können sich auch immer gleichzeitig gegenseitig hemmen. Solche Modalitäten bezeichnen wir als abhängige Modalitäten. Olfaktorische Rezeptoren etwa, die alle die gleichen Outputsignale liefern, gehören zu den abhängigen Modalitäten. Die Wasserströmung in einem Segment unterschied sich jedoch statistisch von der Wasserströmung in einem anderen Segment, so dass die Hemmung nicht vollständig und zu jeder Zeit erfolgte. Daher führte sie nicht zur neuronalen Atrophie. Es gab hinreichend lange Zeiträume, in denen die Signale eines Seitenlinienrezeptors eines Segments nicht von anderen Seitenliniensignalen anderer Segmente gehemmt wurden. Solche Modalitäten bezeichnen wir als unabhängige Modalitäten. Fast alle Rumpfmodalitäten sind unabhängige Modalitäten, denn die abhängigen Modalitäten blieben nur in den Kopfsegmenten erhalten, und dies jeweils nur in einem einzigen Segment. So jedenfalls der Standardfall.
Die sensorischen und motorischen Kerne des vierten Segments entwickelten sich zum Torus semicircularis der Wirbeltiere. In ihnen wurden die Signale des Vestibularsinns mit den Rumpfsignalen abgeglichen und sorgten dadurch für die gewünschte Ausrichtung des Körpers im Schwerefeld der Erde.
Die bereits beschriebene Signalkreuzung der vestibulären Signale ordnen wir dem vierten Segment zu. Hier wechselten die vestibulären Signale die Körperseite. Da dies wahrscheinlich durch einen Wechsel des Körperseitenmarkers gesteuert wurde, wechselten im vierten Segment alle absteigenden und aufsteigenden Axone gleichermaßen zur kontralateralen Seite.
Damit wechselten auch die absteigenden visuellen Signale im vierten Segment zur Gegenseite. Die Augen, die nach dem vierten Segment vor ihrer späteren Rückbildung noch in jedem weiteren Segment folgten, wiesen jedoch keine solche Signalkreuzung auf. Daher stimmten ihre Signale nicht mit denen des zweiten Segments überein. Die laterale Hemmung der Signale aus dem ersten Augenpaar konnte hier nicht wirken. Daher besitzen die höheren segmentierten Tiere, deren Linien zu den Wirbeltieren führen, ein zweites Augenpaar im sechsten Segment. Seine Signalwege sind ungekreuzt. Die Augen der nachfolgenden Segmente fielen jedoch der neuronalen Konkurrenz durch die laterale Hemmung zum Opfer und wurden zurückgebildet.
Damit war der Vestibularsinn in der vierten Etage angesiedelt, die fünfte war dem Seitenliniensinn zugeordnet. Auch hier erwies es sich als günstiger, wenn die Seitenliniensensoren die Muskeln der Gegenseite erregten, so konnte der betreffende Körperteil von der Stelle der Einwirkung wegbewegt werden. Daher wurde auch hier die Signalkreuzung zum Standard und vereinte sich mit der Signalkreuzung des vierten Segments zu einer generellen Signalkreuzung für alle auf- und absteigenden Signale.
Die sechste Etage enthielt ein weiteres Augenpaar, dessen Signalweg ungekreuzt war.
Wir können hier gewiss unwidersprochen annehmen, dass es sich in der Frühzeit der Evolution generell um Facettenaugen gehandelt haben muss. Aus ihnen entwickelten sich im Verlauf der Evolution die Linsenaugen. Doch wie konnte dieser Prozess bei den künftigen Wirbeltieren gestartet worden sein?
Ein Facettenauge (mit hinreichend vielen Facetten) liefert auf der dazugehörigen Rezeptorfläche, die wir durchaus auch als Vorstufe einer Netzhaut bezeichnen können, ein aufrechtes und seitenrichtiges Abbild des Sehraumes. Es gibt keine Linse, die das Bild auf den Kopf stellt und seitenvertauscht abbildet.
Stellen wir uns nun vor, ein fressbares Objekt, eine Beute, befinde sich links im Gesichtsfeld. Es wird beim Facettenauge auf der linken Seite der Retina ein Abbild erzeugen, welches die Sehrezeptoren aktiviert. Diese erzeugen nun Aktionspotentiale.
Vergleichen wir nun den Signalweg des zweiten Segments mit dem des sechsten Segments. In jedem dieser Segmente gab es nach unserer Theorie je zwei Augen. Wir betrachten die Signalwege des linken Auges, denn die Beute ist links zu sehen. Anfangs saßen die Augen seitlich, es gab möglicherweise noch gar kein gemeinsames Sehfeld.
Die visuellen Beutesignale des linken Auges im zweiten Segment aktivierten Neuronen der Klasse 3, die sich im sensorischen Zentrum befanden. Diese projizierten ins motorische Zentrum zu den Neuronen der Klasse 5. Diese wiederum projizierten schwanzwärts zu den Motoneuronen der Rumpfmuskeln. Sie mussten jedoch im fünften Segment die Signalkreuzung durchlaufen, so dass sie sich nun auf der rechten Körperseite befanden. Dadurch führten die visuellen Signale zur Kontraktion der rechten Rumpfmuskulatur, der Tierkörper bog und krümmte sich nach rechts. Der Widerstand des hinteren Rumpfes im Wasser führte dazu, dass auch der Kopf des Tieres nach rechts bewegt wurde. So entfernte sich der Kopf von der Beute. Dies war unzweckmäßig.
Zum Glück gab es aber noch das Augenpaar des sechsten Segments. Dessen Signale mussten keine Signalkreuzung durchlaufen. Daher lösten sie eine motorische Reaktion aus, bei der sich der Rumpf genau nach der anderen Seite krümmte, auf der sich das Futter befand.
So versuchten die Sehsignale aus dem zweiten Segment eine Rumpfkrümmung nach rechts, die aus dem sechsten Segment jedoch eine Rumpfkrümmung nach links. Sie konkurrierten miteinander.
Gewannen die Augensignale aus dem zweiten Segment, so schwamm die Beute fort. Gewannen die Augensignale aus dem sechsten Segment, so konnte man die Beute fressen.
Diejenigen Tiere, bei denen die Augensignale aus dem sechsten Segment den neuronalen Konkurrenzkampf häufiger gewannen, waren klar im Vorteil. So begannen die Augensignale des sechsten Segments, die Augensignale des zweiten Segments zu unterdrücken. Dazu zogen sie auf den Axonen der sensorischen Projektionsneuronen der Klasse 4 kopfwärts, bis sie das sensorische Zentrum des zweiten Segments erreichten. Dort kontaktierten sie hemmende Interneuronen, die nun die visuellen Signale des zweiten Segments zu hemmen begannen.
Damit startete der Prozess der sukzessiven Atrophie des vorderen Augenpaares. Er zog sich über viele Generationen hin. Die nach außen gewölbte Fläche der Facettenaugen im zweiten Segment begann einzufallen und senkte sich in die Tiefe ab, weil die dortigen Strukturen ebenfalls atrophierten. Möglicherweise entstand sogar ein Hohlraum, wo vorher die Augenanlagen waren, denn die Atrophie zeigte bereits Folgen. Es ist nicht schwierig, sich vorzustellen, dass so etwas wie ein primitives Grubenauge entstand, denn die Sehrezeptoren hatten sich ja nicht über Nacht zurückgebildet, weil der Prozess der Atrophie einer Organanlage sich über sehr viele Generationen hinzog.
Die physikalischen Gesetze der Lichtausbreitung führten jedoch dazu, dass Grubenaugen ein seitenverkehrtes, auf dem Kopf stehendes Abbild erzeugen. Durchliefen diese Sehsignale die Signalkreuzung, so wurde diese Verdrehung quasi aufgehoben. Die erste Bildspiegelung um das Bildzentrum erzeugte das Grubenauge, die zweite Spiegelung verursachte die Signalkreuzung. So konnten sich beide neutralisieren. Nun waren die Sehsignale des zweiten Segments wieder brauchbar.
Als sich das Grubenauge weiterentwickelte, indem eine äußere, durchsichtige Schutzschicht entstand, in der sich auch noch eine Sammellinse bildete, war die Abbildungsqualität der Augen im zweiten Segment deutlich besser als die Abbildungsqualität derjenigen im sechsten Segment.
Die Zunahme der Anzahl der Sehrezeptoren in den neuen Linsenaugen des zweiten Segments führte dazu, dass diese den neuronalen Konkurrenzkampf mit den Facettenaugen des sechsten Segments gewannen. Nun hätte man eine vollständige Atrophie der Augen im sechsten Segment erwarten können.
Diese trat jedoch nicht ein. Die relativ einfachen Augen im sechsten Segment hatten in diesem sehr langen Evolutionsprozess als Signallieferanten des optischen Mittelwertkerns im sechsten Segment fungiert. Hier wurde die mittlere Helligkeit ausgewertet und steuerte die Tag- und Nachtaktivität des Tieres. Dieser Steuerungsmechanismus war so wichtig, dass er beibehalten werden musste. Daher übernahmen diese rudimentären Augen die Aufgabe der circadianen Steuerung des optischen Mittelwertkerns. Beim Menschen finden wir dieses optische Steuerzentrum, welches die Aktivität im Tagesablauf steuert, im Nucleus suprachiasmaticus.
Das siebente Segment war – nach dieser Theorie – das Eingangs- und Ausgangssegment des Urhirns dieser Tiere. Hier trafen die Signale aller Rezeptoren aus allen Rumpfsegmenten ein und wurden an die Kopfsegmente übergeben, um mit ihnen in Wechselwirkung zu treten. Der Output wurde wiederum an die Rumpfsegmente übergeben.
Das Urhirn wird also von den Segmenten eins bis sieben gebildet, die wir als Kopfsegmente bezeichnen. Die übrigen Segmente sind die Rumpfsegmente. Möglicherweise wird die künftige Forschung dieses einfache Modell durch weitere Segmente erweitern.
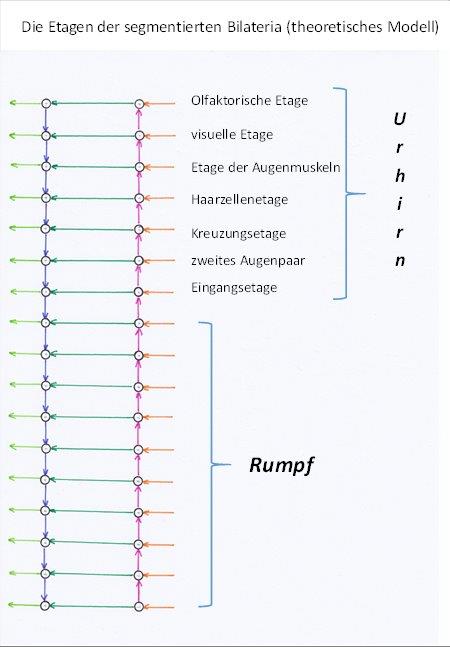
Abbildung 3: Theoretisches Modell des Nervensystems
Die Rumpfsegmente waren – mit Ausnahme des letzten Rumpfsegments, des Knospungssegments – anfangs völlig identisch aufgebaut. Sie besaßen alle die gleichen inneren (und äußeren?) Organe. Doch auch hier ging die Entwicklung weiter.
Organe wurden von den bereits beschriebenen Mittelwertzentren neuronal angesteuert. Doch auch die Mittelwertzentren nahmen an dem segmentüberschreitenden Signalaustausch statt. Jedes Mittelwertzentrum projizierte ebenfalls über Neuronen der Klasse 4 kopfwärts in alle übergeordneten Nachbarsegmente und über Neuronen der Klasse 5 schwanzwärts in alle unteren Nachbarsegmente.
Zwischen diesen Mittelwertsignalen, die jeweils im sensorischen und motorischen Kern eines jeden Segments eintrafen, gab es ebenfalls eine neuronale Konkurrenz. Hemmende Interneuronen sorgten für die laterale Nachbarhemmung.
Dies hatte im Verlauf einer längeren Evolutionszeit zur Folge, dass sich von den vielen gleichartigen Mittelwertkernen, die in jedem Segment vorhanden waren, einige wenige durchsetzten, in anderen Segmenten dagegen atrophierten.
Die Atrophie eines neuronalen Mittelwertkerns in einem Segment hatte zur Folge, dass das von ihm versorgte Organ oder Organsystem in diesem Segment ebenfalls atrophierte, also schrittweise zurückgebildet wurde, da die Steuersignale ausblieben. Gewiss zog sich dieser Prozess über viele Millionen Generationen hin, zumal das Resultat letztlich erblich manifestiert wurde.
Im Endeffekt gab es nicht mehr jedes Organ in jedem Segment, sondern bestimmte Organe nur noch einmal auf jeder Körperseite. Falls dies zur Lebenserhaltung jedoch nicht ausreichte, konnte dieses Organ in mehreren, direkt benachbarten Segmenten erhalten bleiben, in den übrigen Segmenten erfolgte jedoch seine komplette Rückbildung.
Die Tiere auf dieser Entwicklungsstufe waren jedoch noch keine Wirbeltiere. Ihnen fehlte die Wirbelsäule. Aber auch auf neuronalem Gebiet fehlte noch etwas sehr Wichtiges: das Cerebellum.
In einer Monografie über das Gehirn und das Bewusstsein muss der Ursprung der Wirbelsäule nicht unbedingt diskutiert werden. Es gibt zwar die Hypothese, dass die Chordaten sich aus den Manteltieren entwickelten, deren Larven bereits über eine Chorda und ein Neuralrohr verfügen, doch möge dieses Feld anderen überlassen bleiben.
Wie jedoch kam es zur Herausbildung eines Cerebellums? Sämtliche Chordaten verfügen über ein Cerebellum, man könnte sie daher durchaus als Cerebellumtiere bezeichnen.
In meiner Monografie „Gehirntheorie der Wirbeltiere“ habe ich eine Theorie der Entstehung des Cerebellums vorgestellt, die hier in Kurzform wiedergegeben wird.
Ausgangspunkt ist die Bilateralität der hier betrachteten Lebewesen, die als Vorstufe der Wirbeltiere angesehen werden. Bei diesen Tieren standen die linke und die rechte Körperhälfte in einem neuronalen Wettbewerb. Jedes neuronale Zentrum jedes Segments projizierte über Neuronen der Klasse 2 in einen Seitenwechselkern. Dort wurden die Signale auf ein erregendes Projektionsneuron umgeschaltet, welches auf den Körpermarker dieser Körperseite reagierte. Sein Axon zog direkt zum gleichartigen neuronalen Zentrum, welches sich jedoch auf der Gegenseite befand. Dort aktivierte es hemmende Interneuronen, die eine laterale Hemmung der dortigen Neuronen bewirkten. So standen beide Körperhälften in neuronaler Konkurrenz zueinander.
Stärkere Signale auf einer Körperhälfte setzten sich gegen schwächere Signale auf der anderen Körperhälfte durch. Je nachdem, ob Futter rechts oder links gerochen, gesehen oder ertastet wurde, konnte jeweils die richtige Körperseite für die Annäherung an das Futter und für seine Ergreifung sorgen, so dass es gefressen werden konnte. Ebenso konnte Fressfeinden ausgewichen werden, auch wenn wir die Algorithmen hierzu noch nicht immer verstehen.
Die Entwicklung des Cerebellums erfolgte im siebenten Segment. Dieses war gleichzeitig das Eingangssegment des Urhirns unserer Tiere. Das sensorische Zentrum empfing sämtliche sensorischen Signale aus allen Rumpfsegmenten. Dieser Eingangskern spaltete sich bei den Wirbeltieren in zwei Unterkerne auf, die als Nucleus cuneatus und Nucleus gracilis bezeichnet werden. Der Erste erhält die Rezeptorsignale der oberen Rumpfhälfte, der andere die der unteren. Gleichzeitig ist dieses Segment auf der motorischen Seite die Ausgangsetage.
Das motorische Zentrum des siebenten Segments wurde zum Nucleus ruber, dessen Output die Motoneuronen des Rumpfes ansteuert und so für die Bewegungen des Tieres zuständig ist.
Die kontralaterale Hemmung der Gegenseite, die ja in der Anfangszeit in jedem Segment vorkam, wurde dadurch realisiert, dass die Axone der Neuronen der Klasse 2 zum zugeordneten Seitenwechselkern zogen. Dort wurden sie auf ein Seitenwechselneuron umgeschaltet, zogen zur Gegenseite und endeten in dem entsprechenden neuronalen Kern an hemmenden Interneuronen. Der motorische Seitenwechselkern des siebenten Segments wird als Nucleus olivaris bezeichnet. Er empfängt den Output der Nucleus ruber und projiziert in früher Urzeit erregend in hemmende Interneuronen des kontralateralen Nucleus ruber. So wird die kontralaterale Hemmung auf Hirnstammebene in der Frühzeit der Evolution realisiert. Zum Hirnstamm zählen wir die Segmente 2 bis 7, das Segment 1 ist der Cortex.
Die hemmenden Interneuronen des Nucleus ruber, die für diese kontralaterale Hemmung verwendet wurden, trennten sich im Verlauf der Evolution räumlich vom Nucleus ruber und bildeten einen eigenen Kern. Wenn Neuronen sich spezialisieren, finden sie sich zu eigenen Kernen (oder Schichten) zusammen. Dieser neue Kern aus hemmenden Neuronen nahm den erregenden Output des kontralateralen Nucleus olivaris entgegen und schaltete ihn auf den hemmenden Transmitter GABA um. Die Axone seiner Neuronen zogen zum Nucleus ruber und hemmten dort in einer Punkt-zu-Punkt-Abbildung die ihnen zugeordneten Neuronen. So hemmte jedes Neuron des Nucleus ruber das spiegelbildliche Neuron des Nucleus ruber der Gegenseite.
Dieser neue Kern, bestehend aus GABAergen Neuronen, wird von mir in meinen Monografien als Nucleus Purkinje bezeichnen. Seine Neuronen sind die Urform der Purkinjezellen. In der frühen Entstehungsphase des Cerebellums dienten sie der kontralateralen Hemmung des Nucleus ruber der Gegenseite, ihr Input kam vom Olivenkern.
Wenn sich in einem neuronalen Kern eine Neuronenart auf eine spezielle, neue Aufgabe spezialisiert, so trennen sich diese Neuronen oft vom bisherigen Kern und bilden einen eigenen, neuen Kern. Dieser kann als Abkömmling des alten Kerns betrachtet werden. Insofern ist der Nucleus Purkinje ein Abkömmling der hemmenden Interneuronen des Nucleus ruber.
Damit war der erste Schritt in Richtung eines Cerebellums vollzogen. Es fehlte nur noch die Modifizierung des Outputs. Bekanntlich endet der Output der Purkinjezellen in einem Neuronenkern, der aus erregenden Neuronen besteht. Nun können Neuronen mit dem Neurotransmitter Glutamat, wenn sie hemmenden Input erhalten, nur dann einen erregenden Output liefern, wenn sie zusätzlich von externen Signalen erregt werden. Die zugeführte Erregung muss dazu stärker sein als die zugeführte Hemmung. Welcher Kern konnte hier die nötige Erregung liefern?
Im siebenten Segment kann nur der glutamaterge Mittelwertkern infrage. Dieser Kern war die Formatio reticularis (bzw. ihr glutamaterger Anteil). Er erhielt (damals) den Mittelwertinput vom sensorischen und motorischen Kern, also vom Nucleus ruber, vom Nucleus cuneatus und vom Nucleus gracilis. Alle Signale betrafen die Motorik. Darauf hatte sich das siebente Segment spezialisiert.
Mittelwertkerne integrieren möglichst alle verfügbaren Signale. Daher wurden die Signale aus den Purkinjezellen in die Formatio reticularis eingebunden. Hier trat eine neue Art der Signalverarbeitung auf. Die Neuronen des Nucleus reticularis waren (bei Bewegungen) dauererregt, denn sie stellten den Mittelwert der motorischen Erregung dar. Wenn nun eine Purkinjezelle diese Mittelwerterregung hemmte, blieb ein Restsignal übrig. War das Hemmungssignal stark, so war das Restsignal schwach. Wenn aber das Hemmungssignal der Purkinjezelle schwach war, so war das Restsignal im Mittelwertneuron stark.
Dies entsprach der Umkehr der Signalstärke, wir werden dies als Invertierung bezeichnen. Ein starkes Signal wird durch Invertierung in ein schwaches überführt, ein schwaches Signal jedoch in ein starkes. Die Monotonie, also der Signalstärkeverlauf, wird umgekehrt.
Dieser invertierte Output war erregend. Sein motorisches Ziel war nach wie vor der Nucleus ruber. Jedes motorische Signal einer Körperhälfte wurde nun in ein invertiertes Signal umgewandelt und erregte auf der Gegenseite den motorischen Gegenspieler. Dies wird in der neurologischen Fachliteratur als Co-Aktivierung bezeichnet.
Diejenigen Mittelwertneuronen, die den Output der Purkinjezellen empfingen, sonderten sich vom Nucleus reticularis ab und bildeten einen selbständigen, eigenen Kern. Dieser bestand aus glutamatergen Neuronen, die vom Mittelwertkern, dem Nucleus reticularis, eine Mittelwerterregung empfingen. Gleichzeitig wurden sie von den Purkinjezellen gehemmt. Der erregende Output dieses Kerns führte zur inversen Erregung des kontralateralen Nucleus ruber.
Dieser Kern wurde zum ersten Kleinhirnkern des ursprünglichen Cerebellums. Der Purkinjekern stellte nun die Cerebellumrinde dar, in seinem inneren Marklager befand sich der Cerebellumkern, dessen Output den Nucleus ruber mit invertierten Signalen versorgte.
Dieses frühe Cerebellum invertierte die motorischen Signale und führte sie den motorischen Gegenspielern zu. Früher war ein Muskel angespannt, sein Gegenspieler wurde jedoch kontralateral gehemmt. Nun – mit Cerebellum – wurde eine Co-Aktivierung der Gegenspieler erreicht. Der Gegenspielermuskel war nicht mehr völlig erschlafft, sondern verfügte auch über eine gewisse Grundspannung. Dadurch kam Festigkeit in den Körper. So wurde es möglich, dass die ersten Tiere den Lebensraum Wasser verließen. Die Gegenspielermuskeln kompensierten die auftretende Schwerkraftbelastung. Ohne Co-Aktivierung wäre eine Bewegung auf vier Beinen an Land unmöglich gewesen. Insofern ist das Spinocerebellum der Wegbereiter des Landganges der Wassertiere. (Die Entstehung des Vestibulocerebellums ist in meiner vorhergehenden Monografie ausführlich beschrieben worden und kann dort nachgelesen werden.)
Dieses ursprüngliche Cerebellum empfing bereits seit Anbeginn drei Arten von Inputsignalen, jedoch zunächst ausschließlich über die Kletterfaserprojektion von der Olive der Gegenseite:
1. Inputsignale vom Vestibularkern des vierten Segments (Neuronenklasse 5)
2. Motorischen Input der siebenten Etage vom Nucleus ruber (Neuronenklasse 5)
3. Mittelwertinput der siebenten Etage vom Nucleus ruber (Neuronenklasse 6)
Dies führte zur Dreiteilung des Cerebellums in ein Vestibulocerebellum, ein Spinocerebellum und ein Pontocerebellum, ebenso teilte sich der Kleinhirnkern in drei Subkerne, von denen einer sogar zweigeteilt war.
Während die Neuronenklasse 5 den Input für das Vestibulocerebellum und das Spinocerebellum bereitstellte, versorgten die Mittelwertneuronen der Klasse 6 aus dem Nucleus ruber das Pontocerebellum. Dieses war damals noch relativ unbedeutend, da die Anzahl der Mittelwertsignale relativ gering war. Es diente der inversen Aktivierung (Co-Aktivierung) der Mittelwertkerne der Gegenseite.
Erst mit der Herausbildung einer Moosfaserprojektion ins Cerebellum konnte das Pontocerebellum zur Lernmaschine werden, wie schon John C. Eccles vermutete. Dies wird später dargelegt, ist jedoch auch in der vorangegangenen Monografie nachlesbar.
Die Weiterentwicklung des Cerebellums begann mit der Herausbildung einer Moosfaserprojektion. Diese Moosfaserprojektion war möglich, weil die Zellkörper der Nervenzellen im Neuralrohr stets innen angeordnet waren, während außen die Axone kopf- und schwanzwärts verliefen.
Beim Cerebellum empfingen die Purkinjezellen über die Kletterfasern der Olive die Signale der Gegenseite, während die Signale der eigenen Seite in den Axonen der kopfwärts projizierenden Neuronen der Klasse 4 aufwärts zogen. Ebenso befanden sich die sensorischen Neuronen der Klasse 3 und 4 in unmittelbarer räumlicher Nachbarschaft zu den Purkinjeneuronen.
Bereits im Neuralrohr und ebenso im Rückenmark wirkten die Tast- und Schmerzsignale auf die Motoneuronen ein und hemmten diese. Dadurch wurde ein Eigenschutz des Tieres erreicht. Diese hemmende Einwirkung wurde ins Cerebellum übernommen. Realisiert wurde sie dort durch Interneuronen, die wir als Sternzellen und Korbzellen bezeichnen. Die Signale der Tast- und Schmerzrezeptoren erreichten über die Moosfasern die Körnerzellen, deren Axone die Stern- und Korbzellen erregten. Diese hemmten ihrerseits die Purkinjezellen. Diese konnten die Outputneuronen im Kleinhirnkern nicht mehr so stark hemmen, so dass diese stärker feuerten. Die erregenden Signale kamen hierzu vom zugehörigen Mittelwertkern, der Formatio reticularis. Die Outputneuronen des Kleinhirnkerns erregten ihrerseits die kontralateralen Motoneuronen, so dass die Muskeln kontrahierten und den betreffenden Körperteil von der Stelle der Schmerzeinwirkung entfernten.
Wenn nun sehr starke Tast- oder Schmerzsignale der ipsilateralen Seite das Cerebellum erreichten, trafen sie über die Moosfaserprojektion (der Klasse 3) auf die Körnerzellen des Cerebellums und hemmten über die Stern- und Korbzellen die Purkinjezellen. Die stärker gehemmten Purkinjezellen hemmten die Outputneuronen im Cerebellumkern nun deutlich weniger, so dass Letztere auf Grund ihrer von der Formatio reticularis zugeführten Mittelwerterregung nunmehr ein stärkeres Outputsignal lieferten. Da dieser Output die kontralateralen Muskeln erreichte, kontrahierten diese und entfernten den betreffenden Körperteil vom Ort der schmerzhaften Einwirkung. Beispielsweise wurde der Arm weggezogen, wenn eine heiße Herdplatte ihn erwärmte. Somit übernahm das Spinocerebellum den Eigenschutz. Den hatte es zuvor auf Rückenmarksebene gegeben, er wird als Eigenapparat des Rückenmarks bezeichnet. Das Spinocerebellum übernahm im Zuge der Entwicklung der Moosfaserprojektion die Schutzfunktionen des Körpers und den Eigenapparat des Rückenmarks, der dort nun nur noch in Notsituationen vom Rückenmark ausgeübt wurde, da der cerebellare Weg zeitverzögert war.
Später, als die Zahl der Signale der Tiere explosionsartig zunahm, weil eine Unzahl von Rezeptoren gebildet wurde, nahm die Größe des Cerebellums gewaltig zu. Es konnte nicht mehr im Inneren des Neuralrohrs verbleiben. Auch weil die Anzahl der zuführenden Axone der Kletterfasern, die ja stets an der Außenseite des Neuralrohrs zum Cerebellum ziehen mussten, gewaltig zunahm.
Die starke Raumforderung führte zum Aufreißen des Neuralrohrs auf der Höhe des Cerebellums. Durch die geschaffene Öffnung drang das Cerebellum nach außen. Auch die Ventrikelflüssigkeit, die das Innere des hohlen Neuralrohrs füllte, konnte sich nach außen ausbreiten. Seitdem wird das Gehirn auch von außen durch die Ventrikelflüssigkeit umspült. Diese wirkt nun auch als Stoßdämpfer.
Dadurch kehrte sich die Reihenfolge der Neuronenschichten um. Die Neuronen der Klasse 1 befinden sich im Neuralrohr innen an der Ventrikelwand, dann folgen die Neuronenklassen 2 bis 6 in dieser Reihenfolge von innen nach außen. Im Cerebellum und im Cortex suchen die Neuronen der Klasse 1 ebenfalls die Nähe der Ventrikelwand, jedoch wird das Gehirn dort von außen von der Ventrikelflüssigkeit umspült. Daher befinden sich die Neuronen der Klasse 1 im Cortex außen, die der Klasse 6 innen. Damit weist der Cortex folgende Neuronenschichtung von außen nach innen auf:
1. Neuronen der Klasse 1, oben, außen
2. Neuronen der Klasse 2
3. Neuronen der Klasse 3
4. Neuronen der Klasse 4
5. Neuronen der Klasse 5
6. Neuronen der Klasse 6, unten, innen
Wir sollten bedenken, dass eigentlich alle Neuronenkerne eine solche Schichtung aufweisen, auch wenn die Schichtungsreihenfolge in Neuralrohr genau umgekehrt war. Aber die Kopfetagen hatten (am Ende eines Umbauprozesses) sicherlich die gleiche Schichtung wie der Cortex. Vor allem der Thalamus, der ja im zweiten Segment des ursprünglichen Strickleiternervensystems angeordnet war.
Die Neuronen der Klasse 6 sind Mittelwertneuronen. Ihnen verdankt das menschliche Gehirn und ebenso das Wirbeltiergehirn die Fähigkeit zu einer abstrakten Signalanalyse und zur Speicherung von Komplexsignalen. Diese Behauptung wird in dieser Monografie nachgewiesen, ebenso gibt es in meinen früheren Monografien entsprechende Nachweise.
Ohne Mittelwertneuronen wären wir – das wird später beweisen - nicht in der Lage, die Form von Objekten zu erkennen, in unserem Gehirn abzuspeichern und später wiederzuerkennen. Wir hätten nur ein sehr begrenztes Körpergefühl. Wir wissen instinktiv, welche Winkel die verschiedenen Gelenke unseres Körpers bilden, oder welche Lage wir im Schwerefeld der Erde einnehmen. Ebenso sind wir in der Lage, die Bewegung und die Geschwindigkeit von Objekten aller Art einzuschätzen. Dies alles verdanken wir der Existenz von Mittelwertneuronen. Diese magnocellularen Neuronen findet man in vielen Substrukturen des Gehirns, so etwa als Purkinjezellen im Cerebellum, als Pyramidenzellen in der fünften und sechsten Cortexschicht, als GABAerge Striatumneuronen im Basalgangliensystem, aber auch im magnocellularen Anteil des Nucleus ruber oder im Nucleus subthalamicus. Die Funktionsweise der Gehirnmodule, die solche magnocellularen Mittelwertneuronen nutzen, wird im nächsten Kapitel aus informationstheoretischer Sicht analysiert werden.
Ebenso bedeutsam wie die Mittelwertneuronen sind auch die Mittelwertkerne des Gehirns. Ursprünglich in jedem Segment vorhanden, dienen sie der Steuerung wichtiger Lebensfunktionen. Durch die neuronale Konkurrenz zwischen den Signalen aller Segmente untereinander kam es auch hier zu einer Reduktion durch Atrophie, so dass wichtige Mittelwertkerne lediglich in wenigen Segmenten übrigblieben, oft auch nur in einem Kopfsegment. Im Verlauf der Evolution übernahmen die Mittelwertkerne wichtige, völlig neue Aufgaben. Den dopaminergen Mittelwertkernen verdanken wir unser Kurzzeitgedächtnis und die Fähigkeit, Bewegungen und Bewegungsrichtungen zu analysieren. Den glutamatergen Mittelwertkernen verdanken unter anderem wir die Fähigkeit, Komplexsignale zu erlernen. Dies alles wird in dieser Monografie aufgezeigt werden. Wir zählen die wichtigsten Mittelwertkerne des menschlichen Gehirns nachfolgend auf, doch sollten wir bedenken, dass sie alle bereits im Wirbeltiergehirn vorhanden waren.
Mittelwertkerne des Gehirns:
1. Formatio reticularis
2. Substantia nigra pars compacta und Area tegmentalis ventralis
3. Nucleus subthalamicus
4. Septumkerne
5. Raphe-Kerne
6. Locus coeruleus
7. Magnocellulerer Anteil des basalen Kerns der Amygdala
Das Schichtenkonzept des Gehirns wurde auf dem Entwicklungsweg vom niederen Wirbeltier zum Säugetier und vor allem zum Homo sapiens stetig weiterentwickelt. Die Weiterentwicklung war auch eine Folge der Weiterentwicklung der Rezeptoren. Im Verlauf der Evolution wurden immer neue Rezeptorenarten entwickelt. Einige davon waren Neuentwicklungen, zum Beispiel die Muskelspindeln, andere gingen aus bereits vorhandenen Rezeptoren hervor, etwa durch Genverdopplung. Diese neuen Rezeptorenarten ermöglichten es der zugehörigen Modalität, die Umwelt und auch den eigenen Körper immer spezifischer zu analysieren, wodurch die Umwelt- und Körperabbildungen viel aussagekräftiger wurden. Die neuen Rezeptorarten führten zur Aufspaltung der Modalitäten.
Monografie von Dr. rer. nat. Andreas Heinrich Malczan