Signaltheorie der Intelligenz
für das Human Brain Projekt der Europäischen Union
14. Die Zeitanalyse im Gehirn – Übergang zu RNNs
1. Warum Rekurrenz notwendig ist: Statische vs. zeitabhängige Signalverarbeitung
Künstliche neuronale Netze (KNNs) der ersten Generation verarbeiten ausschließlich statische Eingangssignale. Ein Eingabevektor wird durch das Netz propagiert, und das Ergebnis hängt nur von diesem einen Zustand ab. Solche Modelle eignen sich für Aufgaben wie Mustererkennung in Bildern oder Klassifikation einzelner Datenpunkte, berücksichtigen jedoch keine zeitlichen Veränderungen.
Im realen Leben sind Signale jedoch dynamisch. Sie verändern sich entlang einer Zeitachse, und gerade diese Veränderungen bilden oft die entscheidenden Muster. Sprache, Musik, Bewegungen, visuelle Szenen, emotionale Reaktionen und interne Körperzustände sind zeitabhängige Prozesse. Das Gehirn muss daher nicht nur statische Muster erkennen, sondern Sequenzen, Übergänge, Rhythmen und Verläufe.
Um solche zeitlichen Muster zu verarbeiten, entwickelte die KI‑Forschung rekurrente neuronale Netze (RNNs). RNNs besitzen Rückkopplungsschleifen, die frühere Zustände in die aktuelle Verarbeitung einbeziehen. Dadurch können sie zeitliche Komponenten in Signalen abbilden, erlernen und wiedererkennen. Sie sind damit funktional näher an biologischen Systemen, die ebenfalls rekurrente Schleifen nutzen, um zeitliche Informationen zu integrieren.
Das Gehirn arbeitet grundsätzlich rekurrent. Jede Wahrnehmung, jede Entscheidung und jede Bewegung entsteht aus der Integration aktueller Signale mit kurzfristigen und langfristigen Echos vergangener Aktivität. Diese Rekurrenz ist keine technische Ergänzung, sondern ein grundlegendes Prinzip biologischer Intelligenz.
2. Rekurrenz im Gehirn: zwei Systeme, zwei Zeitkonstanten
Das Gehirn verfügt über zwei rekurrente Signalsysteme, die parallel arbeiten und völlig unterschiedliche Zeitkonstanten besitzen. Diese Dualität ist funktional notwendig, weil verschiedene Lern‑ und Verarbeitungsprozesse unterschiedliche Echodauern erfordern.
3. Basalganglien – Kurzzeitechos im Millisekundenbereich
Die Basalganglien erzeugen extrem kurzzeitige rekurrente Signale, typischerweise im Bereich von 10–50 Millisekunden. Diese Echos bestimmen unter anderem das zeitliche Auflösungsvermögen des visuellen Systems. Historisch lag die Wahrnehmungsgrenze bei etwa 18–20 Bildern pro Sekunde (≈ 50 ms Echo). Moderne Generationen zeigen eine feinere zeitliche Auflösung, was sich in der Akzeptanz höherer Bildfrequenzen (60–100 Hz) widerspiegelt. Das Nervensystem hat sich hier innerhalb weniger Generationen funktional angepasst. Die Basalganglien eignen sich damit für schnelle Verstärkungsmodulation, Action Selection und die Sequenzierung motorischer Mikroabläufe – nicht jedoch für langsame Lernprozesse.
4. Papez‑Kreise – Langzeitechos im Minuten‑ bis Stundenbereich
Die rekurrenten limbischen Schleifen des Papez‑Kreises erzeugen deutlich länger anhaltende Aktivitätsmuster. Diese Langzeitechos stabilisieren emotionale, motivationale und episodische Inhalte und liefern die zeitliche Grundlage für Lernprozesse, die Sekunden bis Minuten benötigen. Das Pontocerebellum ist auf solche langen Echos angewiesen, da LTD/LTP‑Mechanismen nicht im Millisekundenbereich arbeiten. Die kurzen Echos der Basalganglien wären hierfür unbrauchbar.
Funktionale Konsequenz
Die Existenz zweier rekurrenter Systeme mit unterschiedlichen Zeitkonstanten ist keine anatomische Kuriosität, sondern eine funktionale Notwendigkeit. Schnelle und langsame Rückkopplungsschleifen erfüllen unterschiedliche Rollen in der Signalverarbeitung und im Lernen. Ein einziges rekurrentes System könnte diese Anforderungen nicht gleichzeitig erfüllen. Dies zwei Systeme (Basalganglien/Papez-Kreise) sind im Gehirn parallelgeschaltet derart, dass ihr Output gemeinsam die rekurrenten Netze erreicht. Zum Lernen mittels LTP/LTD stehen die Signale der Papez-Kreise zur Verfügung, für die schnelle Erkennung die der Basalganglien.
5. Evolution der Rekurrenz: Von Einzelneuronen zu den ersten Differenzschaltungen
Die frühen Wirbeltiergehirne bestanden nicht aus Netzwerken, sondern aus einzelnen Neuronen, die sensorische Signale direkt an Muskeln oder an autonome Mittelwertsysteme weiterleiteten. Diese Architektur erlaubte einfache Reflexe, aber keine Mustererkennung über die Zeit.
Der entscheidende Schritt erfolgte mit der Entwicklung hemmender Neurone. Durch sie konnten erstmals zwei unterschiedlich lange Signalwege zu einem gemeinsamen Outputneuron geführt werden, wobei einer der Wege hemmend wirkte. Der Laufzeitunterschied zwischen den beiden Wegen führte dazu, dass zeitkonstante Signale sich auslöschten, während zeitliche Veränderungen – also Bewegungen – ein Differenzsignal erzeugten.
Diese Schaltung war die Urform des Basalganglien‑Systems: eine frühe rekurrente Struktur, die Bewegungen erkennen konnte, bevor komplexe neuronale Netze existierten.
Der erste Signalweg entstand aus der Notwendigkeit der motorischen Steuerung. Der zweite, hemmende Signalweg entwickelte sich, weil Mittelwertsysteme des autonomen Nervensystems möglichst viel Input benötigen – einschließlich motorischer Signale. Durch die hemmende Rückprojektion zu den Signallieferanten im Cortex entstand dort bereits ein frühes Differenzsignal, das Bewegungen abbildete.
Im weiteren Verlauf der Evolution wurden die hemmenden Interneurone zu Projektionsneuronen, die im motorischen Ausgangskern (Nucleus ruber) die dort vorhandenen motorischen Signale hemmten. Damit entstand die erste explizite Differenzabbildung der Motorik, die später auf digitalisierte Signale erweitert wurde.
6. Rekurrente neuronale Netze - Grundprinzipien
Nach der Herausbildung der biologischen Convolutional Neural Networks (CNNs) im primären Gehirnsystem entstand die Notwendigkeit, nicht nur aktuelle Muster, sondern auch deren zeitliche Abfolge zu analysieren. Dies wurde durch modifizierte CNNs erreicht, die zusätzlich zum aktuellen Input auch zeitverzögerte Signale als Echos empfangen konnten.
Damit entstand eine neue Klasse von neuronalen Netzen: die Recurrent Neural Networks (RNNs)
- Sie arbeiten mit Rückkopplungsschleifen, in denen Signale um einen Zeittakt verzögert erneut als Input verwendet werden.
- Auf diese Weise können RNNs nicht nur räumliche Muster erkennen, sondern auch zeitliche Sequenzen verarbeiten.
- Biologisch entspricht dies der Fähigkeit des Gehirns, Bewegungen, Sprache oder rhythmische Muster zu analysieren und vorherzusagen.
Die RNNs markieren somit den Übergang von der rein räumlichen Mustererkennung der CNNs zur zeitlichen Musteranalyse. Erst auf dieser Grundlage konnte später die nächste Evolutionsstufe entstehen – die Transformer-Netze, die komplexe Divergenz- und Parallelverarbeitungen ermöglichen. Gehirn und KI nutzen die gleichen Algorithmen, unterscheiden sich jedoch in der technischen Realisierung.
Die technische Realisierung von RNNs im Wirbeltiergehirn
Bereits im Kapitel über die Entstehung von CNNs im Wirbeltiergehirn wies ich darauf hin, dass es im Wirbeltiergehirn, insbesondere bei Primaten und beim Homo sapiens insgesamt vier Signalkopien gibt, die aus den cortikalen Signalen hervorgehen:
- Die Projektion in den ipsilateralen Nucleus olivaris und von dort in die Kletterfaserprojektion des kontralateralen Cerebellums (Kletterfaserprojektion)
- Die Projektion über die Nuclei pontis ins Moosfasersysten des kontralateralen Cerebellums
- Die Projektion ins Basalgangliensystem (Matrix und Striosomen)
- Die Projektion in die Papez-Kreise des limbischen Systems.
Für die technische Realisierung der biologischen CNNs ist die Signalkopie in den Papez-Kreisen ausschlaggebend.
1. Die Papez-Kreise als Rotationsspeicher
Die Papez-Kreise des limbischen Systems lassen sich als biologische Rotationsspeicher für neuronale Signale verstehen. Ursprünglich entstanden sie aus der Notwendigkeit, den Duft einer Beute für längere Zeit im Gedächtnis zu behalten – ein evolutionärer Vorteil, da sich Geruchssignale im Wasser des Urozeans zu schnell verflüchtigten.
Im Hippocampus wurden diese Duftsignale in eine höherfrequente Signalfolge transformiert. Die Moosfasern bewirkten dabei eine Zeitverzögerung, indem sie die Signale entlang des Axons streckten. Die Schaffer-Kollateralen zapften diese Moosfasern an und konvergierten auf ein gemeinsames Outputneuron, welches die verteilten Aktionspotentiale integrierte und eine höherfrequente Erregung erzeugte.
Diese Erregung wurde dem zugehörigen Papez-Kreis zugeführt und konnte dort über längere Zeit rotieren.
Signalfluss im Papez‑Kreis
- Hippocampus
- → Fornix
- → Corpora mamillaria (Mammillarkörper)
- → Tractus mamillothalamicus
- → Nucleus anterior thalami
- → Cingulum
- → Cingulärer Cortex / Gyrus cinguli
- → Entorhinaler Cortex / Parahippocampaler Gyrus
- → zurück zum Hippocampus
Die Signalzuführung in die Papez-Kreise erfolgte über drei mögliche Eintrittswege.
Über den cingulären Cortex → Cingulum → Hippocampus
- Assoziationscortices (präfrontal, parietal, temporal) projizieren in den Gyrus cinguli.
- Von dort gelangen die Signale über das Cingulum in den entorhinalen Cortex.
- Der entorhinale Cortex ist das Haupttor zum Hippocampus.
Über den entorhinalen Cortex direkt
- Der entorhinale Cortex erhält massive Projektionen aus nahezu allen Assoziationsarealen.
- Er projiziert direkt in den Hippocampus (Perforant Path).
Über Amygdala / Septum (erweiterter limbischer Kreis)
- Neuere Modelle (MacLean, Yakovlev) erweitern den Papez‑Kreis um Amygdala, Septum und orbitofrontalen Cortex.
Aus den Papez‑Kreisen können die rotierenden Signale an das Cerebellum weitergeleitet werden, wozu sie die folgenden indirekten Projektionswege nutzen:
Über den Hypothalamus → Hirnstamm → Cerebellum
Der Papez‑Kreis enthält die Mammillarkörper, die Teil des Hypothalamus sind. Vom Hypothalamus bestehen Projektionen zu:
- Nucleus reticularis tegmenti pontis (NRTP)
- Pontine Kerne
Diese projizieren massiv in das Cerebellum (v. a. Vermis und Hemisphären).
Über den anterioren Thalamus → Cortex → Pons → Cerebellum
Der Papez‑Kreis sendet über den Nucleus anterior thalami zum cingulären Cortex. Von dort:
- Cingulärer Cortex → motorische / präfrontale Areale
- → Pontine Kerne
- → Cerebellum (Corticopontocerebellärer Trakt)
Dies ist der wichtigste funktionelle Weg.
Fast alle Signale können den Weg in die Papez-Kreise finden. Der Gyrus cinguli besteht aus Millionen Axonen, von denen jedes einem rotierenden Signal zugeordnet ist.
Bei jedem Umlauf erhält das Cerebellum eine Signalkopie, die ins Moosfasersystem eingespeist wird. Diese Signalkopie ist auch dann noch verfügbar, wenn das ursprüngliche Signal bereits endete. So bewahren die Papez-Kreise Vergangenheitssignale für die Gegenwart, indem sie sie in Rotationsschleifen einfangen und dort längere Zeit als Signalechos rotieren lassen.
Das vorhandene neuronale Netz im Cerebellum entwickelte sich durch die Einspeisung der Signalechos zum Recurrent Neural Network (RNN).
2. Hinweis zum Hippocampus
Der Hippocampus dient im hier beschriebenen Zusammenhang lediglich der Erzeugung von Rotationssignalen, die als Echos auch dann noch aktiv sind, wenn das Ursprungssignal bereits endete. Es gibt Hinweise auf Lernvorgänge, die auf die Existenz neuronaler Netze im Hippocampus hindeuten, doch deren genaue Funktionsweise ist bislang nicht vollständig verstanden. Der Autor dieser Monografie verzichtet daher bewusst auf eine detaillierte Beschreibung und beschränkt sich auf die Feststellung, dass der Hippocampus als Quelle zeitverzögerter Signale eine wichtige Rolle für die Rückkopplung in RNNs spielt.
3. Ergänzung: Warum nicht alle Purkinjezellen dasselbe Signal lernen
Eine immer wiederkehrende Frage für das neue RNN lautet: Warum lernen nicht alle Purkinjezellen dasselbe Signal, obwohl die rotierenden Papez-Signale für alle Purkinjezellen dauerhaft zur Verfügung stehen?
Die Antwort liegt in der Topologie der realen neuronalen Netze. Diese verwenden keine Punkt-zu-Punkt-Verbindungen, sondern eine Bustopologie mit lateraler Hemmung im afferenten Signalweg. Bei RNN ist die laterale Hemmung zudem auch auf die Papez-Kreise ausgeweitet. In einer solchen Struktur werden die Signale nicht kontinuierlich allen Empfängern gleichzeitig zur Verfügung gestellt, sondern die Rotation wird durch die Netzarchitektur immer wieder unterbrochen. Dadurch entstehen zeitliche und räumliche Differenzen im Signalfluss, die verhindern, dass alle Purkinjezellen identische Muster erlernen.
Diese Unterbrechungen sind funktional notwendig: Sie schaffen die Grundlage für Diversität im Lernen und ermöglichen die Spezialisierung einzelner Purkinjezellen auf unterschiedliche Muster.
4. Bustopologie und laterale Hemmung in biologischen neuronalen Netzen
Künstliche neuronale Netze (KNN) nutzen eine Ebenentopologie
- Alle Neuronen einer Schicht sind in einer Ebene angeordnet.
- Die Outputneuronen befinden sich in der nächsten Ebene.
- Jedes Neuron der unteren Ebene ist mit jedem Neuron der oberen Ebene verbunden.
- Jeder Input wird an alle Neuronen der Folgeschicht weitergegeben – auch wenn er bereits in einem Knoten erkannt wurde.
- Diese redundante Weitergabe steigert den Energiebedarf erheblich.
Das Gehirn hingegen arbeitet mit einer Bustopologie, die auch in Computern verbreitet ist:
- Alle Inputleitungen einer Schicht verlaufen parallel in einem Datenbus unterhalb der Neuronen.
- Jedes Neuron (Knoten) ist mit jeder Inputleitung verbunden.
- Alle Outputleitungen verlaufen unterhalb der Neuronen ebenfalls parallel.
- Die Neuronen liegen auf einer gedachten waagerechten Linie und bilden eine Knotenkette
- In dieser Bustopologie breiten sich Signale (z. B.) von links nach rechts ohne Richtungswechsel aus.
- In jedem Knoten gibt es einen afferenten Signalweg, der die Inputsignale zuführt, und einen efferenten Signalweg, der die Signale nach der Verarbeitung weiterführt.
- Über hemmende Interneuronen kann jedes Neuron in den efferenten Signalweg eingreifen: Wird ein komplexes Signal (ein Muster) erkannt, werden seine Inputkomponenten aus dem efferenten Signalweg entfernt und stehen nachfolgenden Knoten nicht mehr zur Verfügung.
Die Bustopologie biologischer Netze ermöglicht damit eine ökonomische Signalverarbeitung:
- Erkanntes wird sofort aus dem Datenstrom entfernt.
- Nachfolgende Neuronen müssen nicht dieselben Muster erneut erlernen oder verarbeiten.
- Energieverbrauch und Redundanz werden drastisch reduziert.
Der Unterschied zwischen Ebenentopologie und Bustopologie ist daher fundamental: Während künstliche Netze auf maximale Parallelität setzen, arbeitet das Gehirn mit einer sequentiellen, ökonomischen Signalverteilung, die durch laterale Hemmung zusätzlich abgesichert wird.
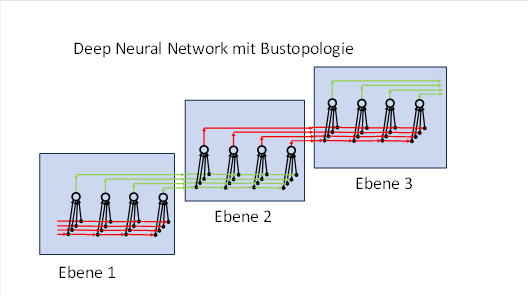
Abbildung: Deep Neural Network mit Bustopologie (Prinzipschaltbild)
5. Signalspezifische, relative Hemmung im efferenten Signalweg
Die Hemmung im efferenten Signalweg beruht ebenfalls auf dem Hebbschen Prinzip und ist signalspezifisch:
- Sie tritt auf, wenn der erkennende Knoten und seine aktiven Inputleitungen gleichzeitig aktiv sind.
- In diesem Fall werden hemmende Interneuronen vom erkennenden Knoten erregt.
- Diese Interneuronen bilden synaptische, hemmende Verbindungen zu den aktiven Inputleitungen (Axonen).
- Solche axo‑axonalen Verbindungen sind im Nervensystem bekannt und ermöglichen die gezielte Unterdrückung derjenigen Signalkomponenten, die das erkannte Komplexsignal bilden, für die nachfolgenden Knoten.
Folgen für die Signalverarbeitung:
- Nur die Elementarsignale des erkannten Komplexsignals werden im efferenten Signalweg gehemmt.
- Nachfolgende Neuronen erhalten diese Inputsignale gar nicht mehr oder nur in stark gedämpfter Form.
- Dadurch entsteht der notwendige Symmetriebruch im Netz: Neuronen spezialisieren sich auf unterschiedliche Signalanteile, statt redundante Muster zu lernen.
- Das Verfahren führt zu einer erheblichen Einsparung an Energie und neuronaler Aktivität
Bedeutung für künstliche neuronale Netze:
-
Die Übernahme der signalspezifischen, relativen Hemmung
zusammen mit der Bustopologie könnte in KNNs enorme Vorteile bringen:
- Reduzierter Energieverbrauch durch gezielte Signalunterdrückung.
- Kürzere Rechenzeiten durch Vermeidung redundanter Verarbeitung.
- Schnelleres Lernen, da nur relevante Signalanteile weitergegeben werden.
6. Spracherwerb mittels RNNs im Gehirn
Die Fähigkeit zum Spracherwerb beruht wesentlich auf der zeitlichen Verarbeitung von Signalsequenzen, die durch RNNs im Gehirn ermöglicht wird. Während CNNs räumliche Muster erkennen, sind RNNs in der Lage, zeitlich aufeinanderfolgende Signale zu speichern, zu vergleichen und zu verknüpfen.
Ab hier raus!
Grundprinzipien:
- Sprache besteht aus Sequenzen von Lauten, die sich über die Zeit entfalten.
- RNNs können durch ihre Rückkopplungsschleifen nicht nur den aktuellen Laut, sondern auch die vorangegangenen Laute berücksichtigen.
- Dadurch entsteht die Fähigkeit, Silben, Wörter und Sätze als zeitliche Muster zu erkennen und zu reproduzieren.
Biologische Umsetzung:
- Die Papez‑Kreise und andere Rückkopplungsschleifen liefern zeitverzögerte Signalkopien, die ins Cerebellum eingespeist werden.
- Dort werden sie mit den aktuellen Inputs kombiniert, sodass Sequenzen von Lauten analysiert werden können.
- Durch laterale Hemmung wird verhindert, dass alle Neuronen dieselben Sequenzen lernen; stattdessen spezialisieren sich einzelne Neuronen auf unterschiedliche Lautfolgen.
- Hebbsches Lernen und synaptische Plastizität (LTP/LTD) sorgen dafür, dass häufige Lautmuster stabil gespeichert werden.
Evolutionäre Bedeutung:
- Ursprünglich diente diese Fähigkeit der Erkennung von Rhythmen und Lautfolgen in der Umwelt.
- Mit der Entwicklung des menschlichen Gehirns wurde sie zur Grundlage für den Spracherwerb.
- Kinder nutzen diese RNN‑Strukturen, um aus den gehörten Lautfolgen die statistisch häufigsten Muster zu extrahieren und zu stabilisieren – ein Prozess, der die Bildung des Wortschatzes ermöglicht.
7. Ergänzung: Sprachverständnis bei niederen Wirbeltieren
Aber auch niedere Wirbeltiere entwickeln ein gewisses Sprachverständnis. Hunde oder Katzen reagieren eindeutig auf ihren Namen – sie „verstehen“ ihn im Sinne einer erlernten Lautsequenz. Papageien können sogar ganze Sätze nachplappern. Dies beweist, dass sie Sprache analysieren können, also über RNN‑Strukturen verfügen, auch wenn sie den Sinn dieser Sätze nicht erfassen.
Damit wird deutlich: Die Fähigkeit zur Verarbeitung zeitlicher Lautfolgen ist evolutionär breit verankert. Beim Menschen wurde sie zur Grundlage des Spracherwerbs und der komplexen Sprachproduktion, während sie bei anderen Spezies in reduzierter Form für Kommunikation und Interaktion genutzt wird.
8. Rückführung erkannter Komplexsignale in die Papez-Kreise
Wenn die Originalsignale und ihre zeitlichen Echos im RNN ausgewertet werden, erreicht der Output (unter anderem) den Cortex und absteigend das Cerebellum. Aber ebenso erreichen sie das limbische System mit seinen Papez-Kreisen, wo sie wieder in Signalechos überführt und an das Cerebellum übergeben werden. Dadurch entstehen tief geschichtete CNNs im Gehirn des Menschen, die Phoneme, Wörter, Sätze und Sprache an sich verarbeiten können.
9. Die Rolle des Gyrus cinguli für biologische RNNs
Im Gyrus cinguli verlaufen beim Menschen Millionen Axone, auf denen die Signale in den zugehörigen Papez-Kreisen rotieren. Sie sind der materielle Beleg für die Existenz riesiger neuronaler Netze mit rekurrenter Signaleinspeisung. Doch auch niedere Wirbeltiere müssen bereits Vorstufen dieser Architektur aufweisen, ist doch ein gewissen Sprachverständnis bei etlichen Tierarten nachgewiesen.
CNNs ohne Cerebellumrinde
Personen, die ohne Cerebellumrinde geboren wurden, jedoch ein gut ausgebildetes limbisches System besitzen, können das neuronale Netz in den Kleinhirnkernen durch Integration der limbischen Signale durchaus zu einem CNN ausbauen, so dass sie zur Mustererkennung fähig sind.
Dritter Hauptsatz für biologische neuronale Netze
Mit der Entstehung des Hippocampus und der Papez‑Schleifen begann die Transformation der cerebellären Netzwerke zu rückgekoppelten neuronalen Netzen. Durch die Einführung eines zeitverzögerten Rückkopplungssignals wurde der frühere Systemzustand als zusätzlicher Eingangswert in die Kleinhirnkerne eingespeist. Die Überlagerung von Vergangenheit und Gegenwart erzeugte komplexe Muster, die die zeitliche Struktur der Signale abbildeten und erstmals prädiktive Funktionen ermöglichten.
Monografie von Dr. rer. nat. Andreas Heinrich Malczan