Signaltheorie der Intelligenz
für das Human Brain Projekt der Europäischen Union
2 Signaltheoretische Grundbegriffe
Ein Elementarsignal ist die kleinste unterscheidbare Einheit eines Signalsystems. Signaltheoretisch entspricht es einem Basisvektor eines geeigneten Vektorraums. Jedes Elementarsignal repräsentiert eine elementare Eigenschaft oder ein elementares Merkmal eines Reizes.
Formal: Ein Elementarsignal ist ein
Vektor
![]() der
kanonischen Basis des Signalraums
der
kanonischen Basis des Signalraums
![]()
Komplexsignale
Ein Komplexsignal ist eine wohlgeordnete Menge gleichzeitig aktiver Elementarsignale. Da wohlgeordnete Mengen eindeutig in Vektorform darstellbar sind, entspricht ein Komplexsignal einer linearen Kombination von Elementarsignalen.
Formal:
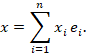
Ein Komplexsignal ist damit ein Punkt im Signalraum, der eine strukturierte Kombination elementarer Merkmale repräsentiert.
Elementarformen und Komplexformen
Eine Elementarform ist die Form, die durch ein einzelnes Elementarsignal repräsentiert wird. Eine Komplexform ist die Form, die durch ein Komplexsignal repräsentiert wird.
Damit gilt:
- Elementarformen sind atomare Muster.
- Komplexformen sind zusammengesetzte Muster, die aus Elementarformen bestehen.
Die Form eines Signals ist seine geometrische Struktur im Vektorraum.
Direktes und inverses System
Ein direktes System bildet Elementarsignale auf Komplexsignale ab. Ein inverses System bildet Komplexsignale wieder auf Elementarsignale zurück.
Formal:
- Direktes System:
![]()
mit
![]() als
Gewichtsmatrix.
als
Gewichtsmatrix.
- Inverses System:
![]()
Im symmetrischen Fall genügt eine einzige
Matrix
![]() ,
da das inverse System durch die Transponierte realisiert wird.
,
da das inverse System durch die Transponierte realisiert wird.
Bedeutung dieser Grundbegriffe
Diese Begriffe erlauben eine präzise signaltheoretische Beschreibung der Vorgänge in mehrschichtigen Netzstrukturen:
- Elementarsignale bilden die atomaren Bausteine.
- Komplexsignale sind strukturierte Kombinationen dieser Bausteine.
- Das direkte System erzeugt Komplexformen aus Elementarformen.
- Das inverse System zerlegt Komplexformen wieder in Elementarformen.
- Die zyklische Anwendung beider Systeme erzeugt einen Wechsel zwischen beiden Darstellungsformen.
Dieser Wechsel bildet die Grundlage für die später beschriebenen Mechanismen der Signalvervollständigung, der Zustandsbildung, der Rekursion und damit der Entstehung von Denken.
Monografie von Dr. rer. nat. Andreas Heinrich Malczan