Gehirntheorie des Menschen
für das Human Brain Projekt der Europäischen Union
ISBN 978-3-00-068559-0
15. DieEvolution des Basalgangliensystems
Erstellt am 12.02.2024 von Dr. Andreas Malczan
Die Hauptaufgabe neuer neuronaler Schaltungen besteht in der Gewinnung neuer Signale.
Die Hauptaufgabe des Basalgangliensystems bestand anfänglich in der Gewinnung von Differenzabbildungen zur Erkennung von Veränderungen der Signalstärke durch die multisensorische Analyse von bewegten Objekten. Erst später, mit der Herausbildung des Cerebellums und der Signaldivergenz und der Einbeziehung des limbischen Systems, ermöglichten die Basalganglien die Verknüpfung von Signalen der Vergangenheit mit Signalen der Gegenwart. Dies war auch eine Voraussetzung zur Verknüpfung von Ursache und Wirkung.
Diese Erkenntnis, hat der Autor erst im letzten Jahr gewonnen, vorher war er der Ansicht, die Bildung von Differenzabbildungen wäre das alleinige Hauptziel des Basalgangliensystems.
Doch die Entwicklung einer Theorie über das Erlernen von Sprache war nur möglich, wenn in den Basalganglien ein Übergang von der Differenzabbildung zur Erzeugung erregender Vergangenheitssignale erfolgte.
Wie ist dies zu verstehen. Nehmen wir eine einfache Ursache: Ein Tier frisst etwas, was nicht bekömmlich ist. Während des Fressens kann es den Geschmack und den Geruch des Gefressenen erlernen. Hierzu dient das Pontocerebellum. Die Ursache kann also erlernt werden.
Etwas später wird dem Tier schlecht. Die damit verbundenen Signale kann das Tier auch erlernen. Die Wirkung wird also ebenfalls abgespeichert.
Doch wenn das Tier die Ursache nicht mit der Wirkung verknüpfen kann, wird es immer wieder das Unbekömmliche fressen und immer wieder wird ihm danach schlecht werden. Falls das Gefressene bei der mehrfachen Aufnahme sogar richtig toxisch ist, wird ein nichtlernendes Tier letztlich daran sterben können.
Das Tier muss also zwei Signalarten verknüpfen können: Vergangenheitssignale mit Gegenwartssignalen. Doch es gibt ein Problem dabei: Vergangenheitssignale sind ohne Weiteres nicht mehr verfügbar. Ihre Lebensdauer ist begrenzt, sie sind halt die Vergangenheit.
Daher musste das Nervensystem der Wirbeltiere eine Weiterentwicklung durchlaufen, bei dem die Vergangenheitssignale in die Gegenwart transportiert wurden, damit sie mit Gegenwartssignalen kombiniert werden konnten.
Dies schaffte das frühe Basalgangliensystem. Es schickte die Signale auf eine Reise, bei der etwas Zeit verging, und in der Zwischenzeit begann bereits die Gegenwart.
Im frühen Urhirn der Wirbeltiere gab es nur analoge Signale, deren Feuerrate die Stärke von sensorischen Signalen repräsentierte. So nahm mit der Geruchsintensität die Feuerrate der Geruchsrezeptoren zu. Ebenso nahm die Feuerrate von Muskelsignalen, beispielsweise der Sehnenorgane, mit der Muskelspannung zu. Alle Signale waren anfangs analog.
Signalstärkeänderungen konnte auch damals die Annäherung oder die Entfernung von Objekten melden, die man roch, ertastete oder sah. Das frühe Striosomensystem der Basalganglien ermöglichte es, die Signalstärkeveränderung zu erkennen.
Alle sensorischen Signale erreichten bereits damals aufsteigend den sensorischen Cortex. Über die Axone der Neuronen der Klasse 3 gelangten die Signale zur motorischen Seite des Cortex und stiegen wieder ab, um über den Nucleus ruber letztlich die Motoneuronen des Rumpfes zu aktivieren. Alle Modalitäten dienten mehr oder weniger der Bewegungssteuerung mit dem Ziel der Futtersuche und der Flucht vor Fressfeinden.
Die absteigenden Cortexsignale gelangten jedoch auch in andere Substrukturen, so beispielsweise in die verschiedenen Mittelwertkerne des Gehirnsystems. Unterhalb des Thalamus gab es das Mittelwertsystem des Nucleus subthalamicus. In der Ausgangsetage des Gehirns befand sich nicht nur der Nucleus ruber, sondern auch das dopaminerge Mittelwertzentrum, welches als Substantia nigra bezeichnet wird. Die abwärts projizierenden Cortexaxone dockten auf ihren Weg auch an diesen zwei Mittelwertstrukturen an und versorgten den Nucleus subthalamicus und die Substantia nigra pars compacta mit erregendem Input. Mittelwertkerne mussten ja zwangsläufig alle verfügbaren Signale in ihre Mittelwertbildung integrieren. Sie selbst dienten unter anderem der Steuerung wichtiger Lebensprozesse.
Nun projizieren Mittelwertkerne fast immer in die Herkunftsstrukturen zurück. Meist dient dies der Aktivierung der sensorischen bzw. motorischen Gebiete, damit beispielsweise die Futtersuche aktiviert werden konnte, bevor der Energiemangel zu groß wurde.
Die Substantia nigra pars compacta projizierte ebenfalls zurück in Richtung Cortex. Da sie den Transmitter Dopamin verwendete, gab es eine dopaminerge Projektion von ihr zum Cortex.
Eine Aktivierung der Cortexneuronen hätte zum neuronalen Aufschaukeln der Signalstärke geführt, da der Cortex wiederum zurückprojizierte. Daher dockten die dopaminergen Axone an den hemmenden Interneuronen des Cortex an und erregten diese. Deren Aufgabe bestand jedoch ursprünglich darin, die benachbarten Cortexneuronen zu hemmen. Diese Hemmungswirkung wurde nun verstärkt. Im Verlauf der Evolution verselbständigten sich diese hemmenden Interneuronen im Cortex und bildeten eine eigene Neuronenschicht unterhalb der Cortexneuronen. Diese Schicht wurde zum Urstriatum. Die darin befindlichen GABAergen Neuronen entwickelten Axonkollateralen, die abwärts projizierten, so wie es die Cortexneuronen auch taten. Sie erreichten im Verlauf der Evolution den Nucleus ruber. Dort konnte die bereits beschriebene Differenzabbildung entstehen. Die hemmenden Signale aus dem Urstriatum überlagerten sich mit den eintreffenden Cortexsignalen, die erregend waren. Die hemmenden Signale hatten jedoch den längeren Umweg über den dopaminergen Mittelwertkern gemacht und waren zeitverzögert, also Vergangenheitssignale.
So entstand im Nucleus ruber eine neue Signalart, die Klasse der zeitsensitiven Differenzsignale. Die hemmenden Striatumneuronen, die ja ursprünglich der Nachbarhemmung dienten, verloren langsam den synaptischen Kontakt mit den Cortexneuronen, die sie ursprünglich gehemmt hatten. Damit war das Urstriatum arbeitsfähig.
Dennoch gab es ein Ziel, welches noch nicht erreicht war. Der Output des Striatums war zwar das Vergangenheitssignal, jedoch war dieses Signal hemmend. Für eine Kombination mit den Gegenwartssignalen war eine erregende Variante erforderlich. Ein erregendes Vergangenheitssignal konnte mit erregenden Gegenwartssignalen auf gemeinsame Outputneuronen konvergieren und so beide sinnvoll miteinander verknüpfen.
Eine Möglichkeit ergab sich durch die Anwendung des Prinzips, möglichst alle Signale in die Mittelwertbildung von Mittelwertzentren einzubeziehen.
Die vom Urstriatum zum Nucleus ruber absteigenden, hemmenden Signale kamen auf ihren Weg direkt am Nucleus subthalamicus vorbei. Im Verlauf der Evolution traten auch diese Signale mit dem Nucleus subthalamicus in Wechselwirkung. Sie dockten an den hemmenden Interneuronen des Nucleus subthalamicus an. So, wie früher die dopaminergen Signale hemmende Interneuronen im Cortex kontaktierten, so nahmen die absteigenden Signale der Striatumneuronen synaptischen Kontakt zu den hemmenden Interneuronen des Nucleus subthalamicus auf und hemmten diese.
Da diese Neuronen jedoch von den Neuronen des Nucleus subthalamicus aktiv erregt wurden, trat nun eine Signalinversion auf. Grund war die relative Hemmung, für eine totale Hemmung waren die Mittelwertsignale des Nucleus subthalamicus zu stark.
Die Fähigkeit hemmender Neuronen zur Kontaktaufnahme mit den hemmenden Interneuronen von Mittelwertkernen und die anschließende Bildung von Projektionsaxonen scheint im Basalgangliensystem eine Grundfähigkeit zu sein, die wiederholt und mehrfach auftritt. Diese Fähigkeit wird uns im Basalgangliensystem noch mehrfach begegnen.
Diejenigen hemmenden Interneuronen des Nucleus subthalamicus, die von den GABAergen Striatumneuronen gehemmt wurden, verselbständigten sich, bildeten einen eigenen Kern und eigene Projektionsaxone. Der von ihnen gebildete Kern war der Globus pallidus, er wird als Globus pallidus interna bezeichnet, weil er sich später in einen internen und einen externen Subkern aufspalten würde. Seine Projektionsneuronen waren GABAerg, also hemmend. Sie hatten vom Nucleus subthalamicus nicht nur die Mittelwerterregung empfangen, sondern auch die Ortsmarker. Daher zogen diese Axone nun wieder aufwärts, wo sie als erste Struktur den Thalamus erreichten. Doch auch im Thalamus gab es bereits Mittelwertneuronen, so wie in den meisten Neuronenstrukturen. Sie befanden sich offenbar auch etwa in der Mitte der damaligen Thalamusstruktur. So gab es im Thalamus eine Art Mittelwertgebiet, einen Subkern mit Mittelwerterregung.
Die aufsteigenden Axone des neu gebildeten Globus pallidus trafen im Thalamus auf diese erregenden Mittelwertneuronen und dockten an ihnen an. Diese Mittelwertneuronen waren vom thalamischen Mittelwert stark erregt, die Signale des Globus pallidus hemmten sie daher nur relativ. Dies entsprach einer Signalinversion.
Die vom Globus pallidus kontaktierten thalamischen Mittelwertneuronen separierten sich im Verlauf der Evolution und bildeten einen eigenen Kern, der neben dem bisherigen Thalamus lag und von Neurologen ebenfalls als Thalamus bezeichnet wird. Dieser neue Thalamuskern erhielt von den Neurologen den Namen Nucleus centromedianus. Seine Neuronen übernahmen von den ursprünglichen Mittelwertneuronen, aus denen sie hervorgegangen waren, deren Ortsmarker und zogen daher aufwärts zum Cortex.
Die Signale des Nucleus centromedianus waren die zweifach invertierten, glutamatergen und zeitverzögerten Cortexsignale. Für die Zeitverzögerung war die Substantia nigra pars compacta verantwortlich. Für die Umschaltung auf GABA waren die Striatumneuronen zuständig. Die erste Inversion fand im Globus pallidus statt, der Output war ebenfalls hemmend. Die zweite Inversion fand im Nucleus centromedianus statt, wo thalamische Mittelwertsignale relativ gehemmt wurden. Der Output war nun erregend und benutzte Glutamat als Transmitter. Er stellte das zeitverzögerte Vergangenheitssignal dar, das nun jedoch durch die Zeitverzögerung in der Gegenwart verfügbar war und mit Gegenwartssignalen kombiniert werden konnte.
Merke:
Die Striosomen des Striatums sind ein Abkömmlinge der hemmenden Interneuronen des Cortex. Der Globus pallidus interna ist ein Abkömmling der hemmenden Interneuronen des Nucleus subthalamicus. Der Nucleus centromedianus des Thalamus ist ein Abkömmling der erregenden Mittelwertneuronen des Thalamus.
Da wir in der bisherigen Analyse stets diejenigen Striatumneuronen betrachtet haben, die durch Dopamin erregt werden, handelt es sich um die Striosomen des Striatums. Daher gilt folgendes:
Output des Striosomensystems
Der Output des Striosomensystems, der wieder den Cortex erreicht, stellt erregende Vergangenheitssignale dar, die durch eine Zeitverzögerung auf ihrem dopaminergen Umweg in die Gegenwart transformiert wurden. Sie können nun mit Gegenwartssignalen kombiniert werden. Diese Signale sind alle feuerratencodiert, stellen also Signale des analogen Gehirnsystems dar.
Doch die Evolution blieb nicht stehen. Schrittweise erfolgte nun eine Signaldivergenz, beginnend im Nucleus olivaris, später auch in anderen Kernen und im Cortex. Sie brachte die Klasse der extremwertcodierten Signale hervor. Auch für diese Signalklasse musste das Basalgangliensystem eine Lösung finden, um sowohl eine Differenzschaltung zur Bewegungserkennung als auch eine erregende Variante der Vergangenheitssignale zu erschaffen. Der Teil des Striatums, der an dieser Aufgabe mitwirkte, war die Matrix. Sie verarbeitete die extremwertcodierten Signale des Gehirns.
Die extremwertcodierten Signale entstanden aus den analogen Signalen durch Signaldivergenz. Zur Verdeutlichung der Signaldivergenz in Nucleus olivaris diene die nachfolgende Grafik des Autors aus der Monografie „Gehirntheorie der Wirbeltiere“:
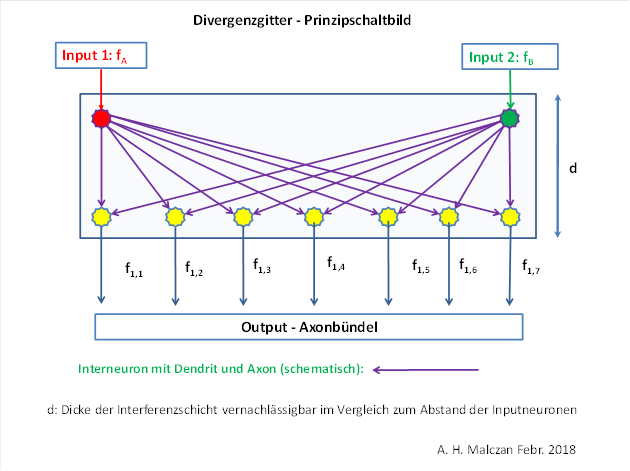
Man sieht, dass durch die Signaldivergenz aus ursprünglich zwei (analogen) Signalen ein ganzer Signalvektor von extremwertcodierten Signalen entsteht, in dem die zwei Ursprungssignale ebenfalls enthalten sind. Im Nucleus olivaris tritt bei linearer Divergenz eine Minimumcodierung auf.
Diese neu hinzugekommenen Signale werden im Basalgangliensystem vom Matrixsystem verarbeitet, welches etwas anders arbeitet als das Striosomensystem.
Zunächst ziehen die minimumcodierten Signale vom Nucleus olivaris als Kletterfasern zum Cerebellum, wo sie die Purkinjezellen stark erregen. Die Purkinjezellen hemmen die dauererregten Kleinhirnkerne, die ihre Dauererregung vom Mittelwertkern der Formatio reticularis empfangen. Die relative Hemmung dieser Dauererregung führt zur ersten Inversion der Signale. Das Spinocerebellum dient als Inversionsschaltung.
Die einmal invertierten Signale sind nun maximumcodiert und erreichen aufsteigend den Cortex. Die Cortexregion liegt im Frontalcortex, der den Cerebellumoutput empfängt.
Der Cortex wiederum projiziert wieder (unter anderem) in den dopaminergen Mittelwertkern, die Substantia nigra pars compacta. Dort werden die Cortexsignale auf Dopamin umgeschaltet und nehmen wieder den Weg zum Striatum.
Hier jedoch ändert sich die Arbeitsweise.
Zwischen diesen neuen Cortexneuronen, die es vor der Signaldivergenz gar nicht gab, entstanden ebenfalls hemmende Interneuronen zur Kontrastverstärkung zwischen den Signalen und zur Unterdrückung schwächerer Signale. Diese Interneuronen waren hemmend und verwendeten GABA als Neurotransmitter.
Im Striatum werden genau diese GABAerge Neuronen kontaktiert. Auch sie dienten ursprünglich auch der Nachbarhemmung. Hierzu kontaktierte jedes Interneuron sehr viele Cortexneuronen und empfing deren Erregung, das Axon des Interneurons hemmte genau ein Cortexneuron. So wirkte die Umgebung dieses Neurons hemmend ein. War die Umgebung relativ stark erregt, so war auch dieses hemmende Interneuron stark erregt.
Wenn nun ein rückkehrendes dopaminerges Axon an ihm andockte und es hemmte, fand eine Signalinversion statt. Der maximumcodierte Cortexoutput wurde durch diese Signalinversion invertiert.
Somit fand im Matrixsystem innerhalb des Striatums bereits die erste Signalinversion der maximumcodierten Cortexsignale des Frontalcortex statt. Die hemmenden Interneuronen erlangten die Fähigkeit, ein eigenes Projektionsaxon zu bilden, welches abwärts projizierte. Der Output der Matrix war hemmend und minimumcodiert.
Die Matrixneuronen behielten ihre synaptischen Kontakte zu den umgebenden Cortexneuronen und vergrößerten ihre dendritischen Felder, um eine stabile Dauererregung vom Cortex zu empfangen, die für die Signalinversion unabdingbar war.
Genauso wie die Striosomenaxone zogen die Matrixaxone in Richtung Nucleus subthalamicus und kontaktierten dort hemmende Interneuronen, so wie es auch die Striosomenaxone taten. Diese Interneuronen sonderten sich ebenfalls ab, bildeten Projektionsaxone und wurden so ebenfalls ein Bestandteil des bereits existierenden Globus pallidus. Allerdings sonderten sich diese Neuronen ebenfalls räumlich ab und bildeten ein eigenes Segment im Globus pallidus. Das ursprüngliche Segment bildete den Globus pallidus interna, während das neue Segment zum Globus pallidus externa wurde, es positionierte sich oberhalb zwischen Striatum und Globus pallidus interna. So entstand eine neue Struktur im Basalgangliensystem.
Der Output des Globus pallidus externa war hemmend, der Transmitter war GABA, die Signale waren nun nochmals invertiert, also maximumcodiert. Technisch unterschieden sie sich von minimumcodierten Signalen des Globus pallidus interna nur durch die Codierungsart. Dieser Kern war genau die Struktur, die nun noch zu durchlaufen war. Daher gab es eine analoge Behandlung dieser Signale.
Die Axone des neuen Globus pallidus externa zogen in Richtung des Globus pallidus interna und generierten dort ebenfalls GABAerge Projektionsneuronen, die wiederum erregenden Mittelwertinput von Nucleus subthalamicus erhielten und somit den Input des Globus pallidus externa invertierten. Damit war der Output des Matrixsystems aus dem Globus pallidus interna wiederum hemmend und minimumcodiert. Die so gewonnenen Signale zogen weiter in Richtung des Thalamus, mussten jedoch zu denjenigen Thalamusstrukturen ziehen, die die ursprünglichen, maximumcodierten Signale vom Cortex erhielten. Auch in diesen Zielgebieten gab es eine verstärkte Mittelwertbildung durch Mittelwertneuronen. Hier dockten die Basalgangliensignale des Matrixsystems, die aus dem Globus pallidus interna kamen, an und nahmen synaptischen Kontakt zu solchen Mittelwertneuronen auf. Die relative Hemmung der Mittelwertsignale entsprach einer erneuten Signalinversion, ihre Outputsignale waren nun erregend und wieder maximumcodiert und entsprachen den zeitverzögerten Cortexsignalen, aus denen sie hervorgegangen waren. Sie stellten jedoch eine neue Signalklasse dar: sie waren erregende Vergangenheitssignale, die der Gegenwart zur Verfügung standen. Über den Umweg der dopaminergen Substantia nigra pars compacta hatten sie die Gegenwart erreicht.
Die zugehörigen Outputneuronen sonderten sich räumlich von den thalamischen Mittelwertneuronen ab und bildeten neue Thalamusstrukturen. So entstanden als neue Thalamusstrukturen der Nucleus ventralis anterior (VA) und die Nuclei ventrales laterales (VL). Sie waren das Ergebnis der evolutionären Weiterentwicklung des Basalgangliensystems unter dem Einfluss der Signaldivergenz im Spinocerebellum.
Nun standen die maximumcodierten Vergangenheitssignale des sekundären Gehirnsystems in ihrer erregenden Form zur Verfügung und konnten mit den Gegenwartssignalen kombiniert werden.
Wir fassen die Arbeitsweise der Basalganglien in der folgenden Übersicht zusammen.
| Striosomen | Matrix | |
|---|---|---|
| Cortexsignale |
analog, erregend Glutamat |
maximumcodiert, erregend, Glutamat |
| Substantia nigra pars compacta | Dopamin, erregend |
Dopamin, hemmend |
| Striatum |
Gaba, Feuerrate bleibt erhalten |
Gaba, Inversionsneuronen, Erregung durch NST, Hemmung durch Dopamin, Output hemmend, minimumcodiert |
| Globus pallidus interna | wird ohne Einwirkung durchquert |
Gaba, Inversionsneuronen, Erregung durch NST, Hemmung durch GABA, Output hemmend, maximumcodiert |
| Globus pallidus externa |
Gaba, Feuerrate wird invertiert, Inversionsneuronen, Erregung durch NST, Hemmung durch GABA, Output hemmend |
Gaba, Inversionsneuronen, Erregung durch NST, Hemmung durch GABA, Output hemmend, minimumcodiert |
| Thalamus, hemmende Variante |
Eine Kopie der Thalamussignale wird vom Globus pallidus externa
in einer Punkt‑zu‑Punkt‑Abbildung gehemmt, zeitsensitive Differenzabbildung |
Eine Kopie der Thalamussignale wird vom Globus pallidus externa
in einer Punkt‑zu‑Punkt‑Abbildung gehemmt, zeitsensitive Differenzabbildung |
| Thalamus, Mittelwertkern |
glutamaterge Inversionsneuronen, Erregung vom Mittelwertkern, Hemmung durch Globus pallidus externa, Output erregend, zeitverschobene Cortexsignale |
glutamaterge Inversionsneuronen, Erregung vom Mittelwertkern, Hemmung durch Globus pallidus externa, Output erregend, zeitverschobene Cortexsignale |
Wie in der Tabelle zu sehen ist, wird sowohl die Bildung der zeitsensitiven Differenzabbildung zur Bewegungserkennung im Basalgangliensystem postuliert als auch die Bildung der erregenden, zeitverschobenen Vergangenheitssignale. Während der Autor in der bisherigen Veröffentlichungen die Bildung der zeitsensitiven Differenzabbildung im Basalgangliensystem erkannt und beschrieben hat, wurde die Bildung der erregenden Vergangenheitssignale erst in der letzten Hälfte des Jahres 2023 von ihm erkannt. Der Grund für die Reifung dieser Einsicht ergab sich, weil das Erlernen von Sprache mittels des lernfähigen Pontocerebellums anders nicht erklärbar war.
Der Autor ist immer noch der Ansicht, dass das Gehirn grundsätzlich Signale verarbeitet, speichert und miteinander kombiniert. Die Übertragung von Vergangenheitssignalen in die Gegenwart ermöglicht dies. Und die einfachste Möglichkeit, ein Signal der Vergangenheit in die Gegenwart zu übertragen, besteht in der Echobildung. Jeder kann das im Gebirge ausprobieren: Ein lauter Ruf breitet sich mit Schallgeschwindigkeit aus und wird von den Bergwänden reflektiert. Während seiner Ausbreitung läuft die Uhr weiter. So trifft der Schall beim Hörer zu einem viel späteren Zeitpunkt ein als er erzeugt wurde, ist also ein Signal aus der (gerade verflossenen) Vergangenheit.
Die Basalganglien nutzen die geringere Ausbreitungsgeschwindigkeit und den längeren Signalweg über den dopaminergen Mittelwertkern, um eine solche Zeitverzögerung des Signals zu erreichen.
Nun müssen wir uns noch mit der Größe der Zeitverzögerung befassen. Diese ist so gering, dass es nun noch weiterer Subsysteme des Gehirns bedarf.
Die Zeitverzögerung im Basalgangliensystem liegt deutlich unter dem Sekundenbereich, der Autor schätzt sie auf etwa 25 bis 50 Millisekunden. Das bedeutet, dass das Echo noch etwa 25 bis 50 Millisekunden nach dem Ende des Ausgangssignals aktiv ist. Diese Zeit ist zu kurz, um in Pontocerebellum irgendwelche Lernvorgänge mittels LTP und LTD zu bewirken.
Daher benötigt das Gehirn eine Möglichkeit, diese Signaldauer deutlich zu verlängern, so dass sie mindestens eine Sekunde oder mehr beträgt. Je länger die Echodauer eines Signals ist, um so größer ist die Zeitspanne, in der es mit anderen, später aktivierten Signalen kombiniert werden kann.
Zur Lösung dieses Problems konnte eine Struktur beitragen, die es bereits seit Urzeiten im Wirbeltiergehirn gab. Diese Struktur war der Papez-Kreis.
Jedes Signal, welches in den Papez-Kreis eingespeist wird, rotiert dauerhaft in einer geschlossenen Schleife mit folgenden Zwischenstationen:
·Hippocampus
·Corpus mammilare
·Nucleus anteriores thalamii
·Gyrus cinguli
·Area entorhinalis
·Gyrus parahippocampalis
·Tractus perforans
·Hippocampus
Die Signalrotation in der limbischen Schleife, wie der Papez-Kreis auch genannt wird, könnte theoretisch ewig stattfinden. Praktisch gibt es jedoch den Zugriff über Stoppsignale, die an verschiedenen Stellen der Schleife hemmende Interneuronen erregen können, die ihrerseits die Signalrotation stoppen. Die innere Uhr im Gehirn im Nucleus suprachiasmaticus kann solche Stoppsignale an das limbische System liefern. Aber auch andere Regionen können dies. Insbesondere kann zwischen den limbischen Signalen eine neuronale Konkurrenz auftreten, so dass ein Signal andere Signale unterdrücken kann. Dies wird uns im nächsten Kapitel begegnen.
Das limbische System kann die zeitlich recht kurzen Signalechos des Basalgangliensystems deutlich verlängern, so dass ihre Lebensdauer weit über den Sekunden- oder gar Minutenbereich hinausreichen kann. Dazu ist es lediglich erforderlich, dass diese Signale dem limbischen System als Input zugeführt werden. Wenn sie dort in Schleifen rotieren, können sie bei jedem Signalumlauf als Output über die Brückenkerne und die Kletterfasern das Pontocerebellum erreichen. Nun kann das Pontocerebellum diese Vergangenheitssignale dazu verwenden, mit Gegenwartssignalen kombiniert zu werden. Wie dies geht, wird der Autor beispielhaft im nächsten Kapitel erklären. In diesem Kapitel wird erklärt, welche Algorithmen im Gehirn in welchen Subsystemen das Erlernen der gesprochenen Sprache ermöglichen. Das Erlernen der geschriebenen Sprache wird voraussichtlich in einem späteren Kapitel behandelt.
Monografie von Dr. rer. nat. Andreas Heinrich Malczan