Gehirntheorie des Menschen
für das Human Brain Projekt der Europäischen Union
ISBN 978-3-00-068559-0
Divergenzmodule des Gehirns
In einem Divergenzmodul kommt es zur Überlagerung von Signalen verschiedener Herkunft. Sie treffen in unterschiedlichen Inputschichten ein, zwischen denen Outputschichten liegen. Durch Signaldämpfung kommt es zur Extremwertcodierung des Outputs. Wie kam es dazu?
Der letzte wichtige Schritt auf dem Wege des Wirbeltiergehirns in Richtung Primatenhirn erfolgte ebenfalls schrittweise und begann schleichend in früher Urzeit.
Neuronen waren störungsanfällig, sie konnten aus vielerlei Gründen fehlerhaft funktionieren oder sogar absterben. Da war es günstig, wenn Signale über mehrere, parallele Signalwege übertragen wurden. Zwei Neuronen, deren Axone das gleiche Signal übertrugen, verdoppelten die Signalsicherheit.
Verteilte man ein Signal gleich auf mehrere Neuronen, dann stieg die Zuverlässigkeit weiter an. Dann konnten sogar mehrere Neuronen ausfallen, ohne dass der Körper einer wichtigen Funktion beraubt wurde.
Natürlich musste bei der Zielstruktur, die dieses Signal empfangen sollte, die Signaldivergenz rückgängig gemacht werden. So finden wir in vielen Strukturen große, magnocellulare Neuronen mit großem Dendritenbaum, die das zuvor auf mehrere oder viele Axone verteilte Signal wieder empfangen und die Signaldivergenz rückgängig machten. Solche Konvergenzneuronen finden wir etwa im magnocellularen Teil des Nucleus ruber oder als Betzsche Pyramidenzellen im Cortex, ebenso im Striatum, wo sie die Striosomen bilden. So konnte die Signaldivergenz durch Konvergenzneuronen rückgängig gemacht werden.
Im Verlauf der Evolution wurde dort, wo genügend Platz vorhanden war oder wo er sich leicht schaffen ließ, eine redundante Signalübertragung zum Standard. Bereits im Neuralrohr wird ein Signal oft auf mehrere Neuronen verteilt.
Ein Neuronenkern im menschlichen Gehirn, der Nucleus olivaris, weist eine besonders starke Signaldivergenz auf. Hier wird jedes Inputsignal auf eine sehr große Anzahl von Outputneuronen verteilt. Dadurch bläht sich dieser Kern extrem auf.
Da sein Output das Cerebellum erreicht, wächst auch dort die Anzahl der Purkinjezellen fast ins Unermessliche. Die Cerebellumrinde dehnt sich extrem aus. Durch Platzmangel wird die Oberfläche faltig, damit sie sich noch weiter vergrößern kann.
Aber auch der Cortex und alle seine Cortexlappen zeigen diese Signaldivergenz. Die Anzahl der cortikalen Inputneuronen ist extrem klein im Verhältnis zu den cortikalen Outputneuronen.
Die Höhe der Evolutionsstufe einer Art lässt sich deutlich mit der Größenzunahme des Nucleus olivaris, des Cerebellums und des Cortex in Zusammenhang bringen, die vorwiegen, jedoch nicht ausschließlich durch die Signaldivergenz entsteht.
Diese Tendenz nimmt beim menschlichen Cortex einen extremen Grad an. So wird sogar die Cortexoberfläche in Falten gelegt, um im vorhandenen Volumen eine maximal große Anzahl von Outputneuronen unterzubringen.
Hier muss ich als Autor dieser Monografie den Anhängern und den Planern des Human Brain Projekts den Vorwurf machen, der Frage nach dem Grund dieser gewaltigen Zunahme der Neuronenanzahl im Nucleus olivaris, im Cerebellum und im Cortex (und anderen Strukturen) überhaupt nicht auf den Grund gegangen zu sein und auch die Folgen dieser Entwicklung zu vernachlässigen.
Diese Forscher stellten das unbewiesene Dogma auf, Intelligenz und Denken sowie die gesamte Funktionsweise des Gehirns sei allein mit dem Studium der synaptischen Verbindungen zu erklären. Einen Beweis für die Richtigkeit dieses Dogmas konnte bisher nicht erbracht werden.
Es ist etwa so, als würde man behaupten, die Gesetze der Wärmeausbreitung entlang eines Kupferdrahtes seien durch das Studium der Geschwindigkeit der einzelnen Kupferatome erkundbar, daher müsse die Geschwindigkeit eines jeden Atoms zunächst exakt ermittelt werden.
Natürlich hängt die Temperatur eines Kupferdrahtes von der Geschwindigkeit der Kupferatome ab, die als Brownsche Bewegung bezeichnet wird, aber das Resultat ist ein statistisches. Der Mittelwert der Geschwindigkeit der Atombewegungen gibt die Temperatur vor. Statistisch kann man vom konkreten Atom abstrahieren, und in den Formeln zur Temperaturverteilung kommen keine Atome mehr vor.
Und die Temperaturverteilung entlang des Kupferdrahtes hängt auch zusätzlich von der Entfernung zur Wärmequelle ab. Hier bedarf es der Lösung einer Differentialgleichung unter Berücksichtigung der Randwerte.
Warum komme ich mit diesem einfachen Beispiel der Wärmeausbreitung in einem Kupferdraht?
Weil zwischen der Ausbreitung von Wärme in Stoffen und der Ausbreitung einer neuronalen Erregung möglicherweise eine große Analogie besteht.
Stellt man sich die Ausbreitung der Erregung ebenso vor wie die Ausbreitung von Schall, Wärme oder elektrischen Ladungen in Form von Ionenwolken vor, so abstrahiert man von der Existenz von Atomen, Molekülen oder auch Neuronen. Man betrachtet nur noch den Ort der Signaleinspeisung, den Ort des Signalempfangs und den Abstand zwischen beiden. Man unterstellt eine abstandsabhängige Dämpfung der Schallintensität, der Wärmeübertragung oder der neuronalen Erregung. Nun bestimmt man in den theoretisch hergeleiteten Formeln die Übertragungsparameter und bestätigt sie experimentell.
Neuronen sind in diesem Modell überflüssig.
Es ist nicht hinnehmbar, dass die Experten des Human Brain Projekts, insbesondere die Mathematiker und Physiker unter ihnen, einfache Naturgesetze, die für die Ausbreitung von neuronalen Erregungen gelten, einfach ignorieren, obwohl sie inzwischen selbst in der (unter Wissenschaftlern oft abschätzig bewerteten Wikipedia) zu finden sind. So findet man in der deutschen Wikipedia unter dem Suchbegriff -Aktionspotential- den bemerkenswerten Satz:
-1952 legten Alan Lloyd Hodgkin und Andrew Fielding Huxley ein mathematisches Modell [9] vor, das die Entstehung des Aktionspotentials im Riesenaxon des Tintenfisches durch das Wechselspiel verschiedener Ionenkanäle erklärt und unter dem Namen Hodgkin-Huxley-Modell berühmt wurde. Für diese Entdeckung erhielten die beiden Forscher zusammen mit John Eccles 1963 den Nobelpreis für Medizin.-
Ebenso findet man in der Wikipedia die komplette Herleitung der Kabelgleichung für die Erregungsausbreitung auf nicht myelinisierten Axonen.
Allen Neurowissenschaftlern ist die Tatsache bekannt, dass die Großhirnrinde zur sogenannten grauen Substanz gehört und die in ihr verlaufenden Axone (meist) von keiner Myelinschicht umgeben sind. Ausnahmen gibt es in einigen Bereichen der visuellen Rinde, wo makroskopisch erkennbare weiße Streifen innerhalb der grauen Substanz auftreten (Gennari-Streifen oder Vicq-d'Azyr-Streifen). Deshalb wird dieses Cortexareal auch als Area striata (deutsch: gestreifter Bereich) bezeichnet. Offenbar wird hier eine wichtige Information über schnellleitende, myelinisierte Axone in benachbarte Cortexfelder weitergeleitet. Diese Information muss mit der Bewegungserkennung zu tun haben, weil sie ihre Auswertungsgebiete zeitnah erreichen muss.
Die Existenz der grauen Substanz in der Hirnrinde gibt uns zusammen mit den Naturgesetzen der neuronalen Erregungsausbreitung, insbesondere der Kabelgleichung für marklose Fasern die entscheidenden Hinweise, die Signalverarbeitung in der Cortexrinde und in allen übrigen Kernen mit marklosen Neuronen wissenschaftlich zu analysieren.
Insofern muss den im Human Brain Projekt beteiligten Forschern und vor allem den für dieses Projekt verantwortlichen Koordinatoren und Planern den Vorwurf machen, geltende Naturgesetze fahrlässig oder gar vorsätzlich nicht beachtet oder sogar mit Füßen getreten zu haben. Das völlige Fehlen irgendwelcher Reaktionen auf meine bisherigen Veröffentlichungen in meiner Monografie -Gehirntheorie der Wirbeltiere für das Human Brain Projekt der Europäischen Union- zeigt die Ignoranz der Verantwortlichen.
Die Kabelgleichung für die Ausbreitung von neuronalen Erregungen entlang eines Axons gibt uns die Richtung für eine neue Sichtweise vor. Es zeigt sich, dass nur der Abstand des Zielortes vom Ort der Signaleinspeisung zählt. Die an der Signalausbreitung beteiligten Unmengen von neuronalen Synapsen, die in die Zehntausend oder Hunderttausend gehen können, sind prinzipiell belanglos!
Breitet sich eine neuronale Erregung entlang eines Axons aus, dessen Durchmesser als konstant angenommen wird, so gilt für die Erregung im Abstand r vom Axonanfang die Gleichung
![]() ,
,
hierbei ist λ die sogenannte Längskonstante des Neurons. Der Indes l in El kennzeichnet, dass hier die lineare Ausbreitung entlang eines Axons vorliegt.
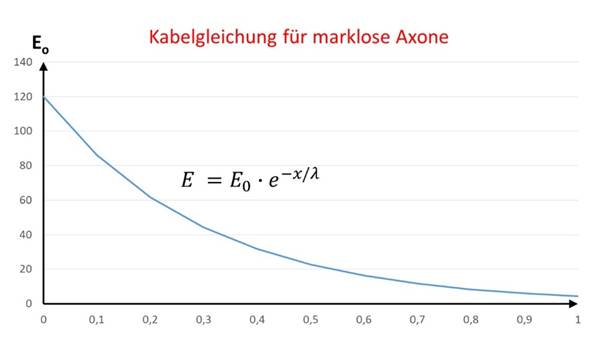
Abbildung 6: Kabelgleichung für marklose Fasern
Oft breitet sich die Erregung jedoch in der Fläche aus. Ein cortikales Inputneuron erregt eine Unzahl von Outputneuronen der näheren oder ferneren Umgebung in der Cortexfläche. Dazu bildet es entweder einen weitverzweigten Dendritenbaum oder nutzt Interneuronen für die Signalverteilung in der Fläche. Hier sind wiederum fast unzählig viele Synapsen beteiligt, die man jedoch völlig vernachlässigen kann, da hier nur die Entfernung vom Einspeisungspunkt und die Längskonstante des Inputneurons in die Gleichung eingehen. Dies ist zulässig, weil die Cortexrinde zur grauen Substanz zählt.
In diesem Fall muss die Kabelgleichung für eine Erregungsausbreitung in der Ebene präzisiert werden, im negativen Exponenten geht anstelle des Abstandes r das Quadrat r2 des Radius des Kreises ein, in dessen Fläche sich die Outputneuronen befinden.
mit
![]() .
.
Der Grund liegt darin, dass die Ionenwolke, die letztlich für den Potentialunterschied zwischen Zellinnerem und dem extrazellularen Raum verantwortlich ist, sich nicht linear entlang eines einzelnen Axons ausbreitet, sondern sich in der Ebene verteilt. Natürlich bedarf es dazu der Axone, aber diese verzweigen sich extrem und versorgen so die Zielneuronen in der Umgebung.
Und es ist natürlich fraglich, ob die Kabelgleichung für marklose Fasern in ihrer bisherigen auf den Cortex angewendet werden kann. Ihre experimentelle Überprüfung erfolgte ursprünglich am Riesenaxon des Tintenfisches. Die Axone der Neuronen und Interneuronen im Cortex, aber auch in anderen grauen Kernen sind deutlich dünner, viel verzweigter und besitzen teils eine räumliche Ausdehnung. Daher habe ich in meinen Monografien für diese Neuronenarten eine exponentielle Dämpfung angenommen, die quadratisch vom Abstand abhängt. Diese Art der Erregungsausbreitung ist im nachfolgenden Diagramm symbolisch dargestellt.

Abbildung 7: Erregungsausbreitung in der Ebene - seitlich gesehen
Von oben gesehen ergibt sich die folgende Ansicht:
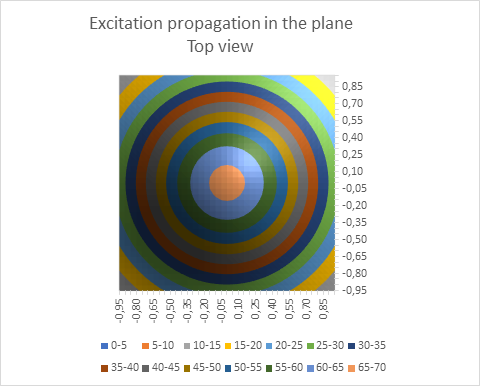
Abbildung 8: Erregungsausbreitung in der Ebene - von oben gesehen
Oft breitet sich die neuronale Erregung im Raum aus - die Cortexschichten sind ja nicht unendlich dünn. In diesem Falle könnte man annehmen, dass der Radius mit der dritten Potenz in die Kabelgleichung eingehen würde:
![]() .
.
Einen solchen Ansatz habe ich verworfen. Dazu müssten die Anzahl der Axomverzweigungen gegen Unendlich gehen und sich im Raum quasi kontinuierlich verteilen.
Dies ist jedoch nicht so. Einfacher vorstellbar ist, dass man sich zunächst vorstellt, die Axone verzweigen sich in der Ebene baumartig. Hier wäre eine quadratische Abhängigkeit zu erwarten, da sich die Erregung annähernd in der Fläche verteilt.
Anschließend stellt man sich vor, die vorhandenen Axone könnten einfach so verbogen werden, dass sie baumartig räumlich verzweigt würden. Durch die Verbiegung würde ihre Übertragungskennlinie ja nicht verändert.
Damit haben wir eine neue Formel für die neuronale Erregung für die Ausbreitung in der Ebene und im Raum.
Es ist sicherlich einfach, sich vorzustellen, dass die Feuerrate f eines Neurons, welches sich an der Stelle x oder im Abstand r vom Signallieferanten befindet und dessen Erregung f0 unter Beachtung einer Signaldämpfung empfängt, ebenfalls von dieser Entfernung abhängt. Auch hier geht der Abstand mit der zweiten Potenz in die Formel ein, und anstelle der Eingangserregung E steht die Feuerrate f dieses Signallieferanten.
Dann gilt in erster Näherung für die Feuerrate f
Hierbei gilt für den Radius r in der Ebene die Gleichung
![]() .
.
Im Raum gilt für den Radius r die Gleichung
![]() .
.
Im Allgemeinen wird nicht ein einzelnes Inputneurone seine Erregung zum Cortex oder in eine andere Struktur leiten, sondern es sind immer mehrere benachbarte Inputneuronen im Cortex, die aktiv sind. Dann überlagern sich die Erregungsfelder der verschiedenen Inputneuronen additiv, es entsteht eine Summenerregung.
Nun kann man nicht einfach die Kabelgleichung des Hodgkin-Huxley-Modell
![]() (Hodgkin-Huxley-Modell)
(Hodgkin-Huxley-Modell)
durch die Gleichung
![]() (in dieser Monografie verwendet)
(in dieser Monografie verwendet)
ersetzen, weil man diese aus physikalischer Sicht für cortikale Neuronen für besser geeignet hält. Es müssen weitere handfeste Gründe dafür sprechen.
Ein handfester Grund ist das Auftreten von Maximalwerten der neuronalen Erregung, die bei Veränderung von Untersuchungsparametern umherwandern. Bei der Überlagerung von neuronalen Erregungen können veränderliche Maximalwerte bei Anwendung der Hodgkin-Huxley-Gleichung nicht auftreten. Es sind nur ortsveränderliche Minima möglich.
Wir diskutieren die Erregungsausbreitung und die Entstehung von Erregungsmaxima aus mathematischer Sicht.
Wir beschränken uns in dieser Monografie auf den Fall der Erregungsausbreitung in einer Ebene, dies entspricht näherungsweise der Erregungsausbreitung in einer relativ dünnen Neuronenschicht in der Cortexrinde. Die Inputneuronen liegen in der vierten Cortexschicht, hier breitet sich die neuronale Erregung (vereinfacht) in der Fläche aus.
Die Erregungsfunktion ist in einem gewissen Kreis um das Inputneuron streng konkav und besitzt an der Stelle des Inputneurons ein Maximum. Dies zeigt die Berechnung der Hessematrix.
Ausgangspunkt ist Erregungsfunktion f mit der Funktionsgleichung für die Ebene. Die Funktion f ist zweimal stetig differenzierbar.
![]()
Die Hessematrix H ist definiert über die zweiten partiellen Ableitungen:
![]() .
.
Die Berechnung der partiellen Ableitungen ergibt die Hessematrix:
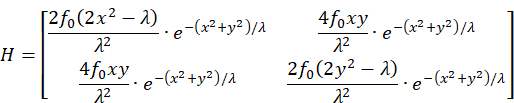
Die Hessematrix ist genau dann negativ definit und die Funktion f streng konkav, wenn alle geraden Hauptminoren größer als Null sind, jedoch alle ungeraden Hautpminoren kleiner als Null sind.
Der erste Hautpminor ist gleich fxx und ist negativ genau dann, wenn die Bedingung
![]()
erfüllt ist. Dies trifft zu für alle Werte x, die die Ungleichung

erfüllen.
Der zweite Hauptminor der Hessematrix ist gleich H und muss größer als Null sein. Daher muss die Gleichung
![]()
erfüllt sein. Dies bedeutet
![]() .
.
Unter Beachtung der Tatsache, dass die Längskonstante λ > 0 ist, der Feuerratenfaktor fo > 0 ist, und die Exponentialfunktion ebenso immer größer als Null ist, bleibt als Bedingung nur noch
![]() .
.
Dies ist unter Beachtung von
![]()
genau dann erfüllt, wenn die Ungleichung
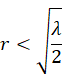
erfüllt ist.
Dann ist die erste Ungleichung ebenfalls automatisch erfüllt.
Somit ist die Erregungsfunktion
![]()
genau dann streng konvex, wenn der Radiusvektor r die Bedingung
erfüllt. Innerhalb des Kreises mit diesem Radius r ist diese Funktion streng konvex und besitzt innerhalb dieses Kreises das einzige, globale Maximum.
Eine Verschiebung der Funktion an einen anderen Punkt führt zur Verschiebung des Kreises, in dem die verschobene Funktion strikt konkav ist. Das bedeutet Folgendes:
Die verschobene Funktion
![]()
Ist streng konkav und besitzt ein globales Maximum in dem Gebiet, welches der Bedingung
![]()
genügt.
Jedes im Punkt x0 und y0 befindliche Inputneuron mit der Feuerrate f0 besitzt genau diese streng konkave Erregungsfunktion.
Eine additive Überlagerung der Erregungen von m Inputneuronen N1, N2, -, Nm, die sich jeweils in den Punkten P1(x1;y1), P2(x2;y2), -, Pm(xm;ym) befinden, führt auf die nachfolgende Gleichung für die Gesamterregungsfunktion f:
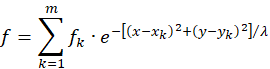
Überlagern sich derartige Erregungsfunktionen fk, die an verschiedenen Stellen im Koordinatensystem angeordnet sind, wobei sich die Kreise, in denen sie strikt konkav sind, sich ebenfalls überlagern, so ist die Existenz eines gemeinsamen globalen Maximums in dieser Überlagerungsfläche gesichert. Für hinreichend große Werte von λ (zum Beispiel für λ = 9) besitzen beliebig viele Überlagerungen solcher Erregungsfunktionen immer genau ein globales Maximum, wenn sich die signalliefernden Inputneuronen beliebig in einem Quadrat mit der Seitenlänge 2 verteilen, wenn der Ursprung des Quadrats im Koordinatenursprung liegt und die Quadratseiten beispielsweise parallel zu den Koordinatenachsen ausgerichtet sind. Die strikte Konvexität einer solchen Überlagerung ergibt sich dann automatisch.
Die einzigen Parameter, die in der Gesamterregung f vorkommen, sind die Feuerraten f1, f2, -, fm der m Inputneuronen Dies können zum Beispiel die Signale von Muskelspindeln eines Gelenks sein, ebenso die Signale der zugehörigen Sehnenrezeptoren, aber auch die Signale benachbarter magnocellularer visueller Projektionsneuronen. Fast jede Modalität kann ihre Signale in solche Divergenzmodule schicken. Erkennbar sind solche Divergenzschaltungen eindeutig daran, dass die Anzahl der Inputneuronen im Vergleich zur Anzahl der Outputneuronen bedeutend kleiner ist. Ein weiteres Merkmal ist das Auftreten von Erregungsmaxima, die umherwandern, wenn die Inputfeuerraten sich verändern.
Ein Hinweis noch zu den Orientierungssäulen im primären visuellen Cortex. Sie reagieren selektiv auf den Anstellwinkel einer geneigten Geraden. Ein Neuron in einer solchen Orientierungssäule reagiert mit einer maximalen Erregung, wenn der Anstellwinkel der Geraden einen bestimmten Wert φ annimmt. Bei anderen Winkeln nimmt die Erregung ab. Es ist mir völlig unverständlich, wie man versucht, dieses Verhalten des Neurons über Lernvorgänge in neuronalen Netzen zu erklären. Warum so umständlich, wenn eine einfache Lösung auf der Hand liegt? Eine Lösung, die einfache Gesetzmäßigkeiten von periodischen Funktionen und ihren Linearkombinationen nutzt.
Wenn eine dunkle Gerade das kreisförmige rezeptive Feld einer magnocellularen Ganglienzelle schneidet, die auf dunkle Objekte reagiert, so ist die Feuerrate dieser Zelle eine streng monotone Funktion der Sehnenlänge.
Rotiert die Gerade um einen Punkt innerhalb des rezeptiven Feldes, so ist die Sehnenlänge und damit auch die Feuerrate eine periodische Funktion des Anstellwinkels φ der Geraden. Die Periode dieser Funktion ist gleich π.
Sind vier solcher Ganglienzellen mit kreisförmigen rezeptiven Feldern derart angeordnet, dass sich ihre rezeptiven Felder überlappen und ein gemeinsames Teilgebiet besitzen, und rotiert eine dunkle Gerade um einen festen Punkt in diesem gemeinsamen Gebiet, so ist die Feuerrate einer jeden Ganglienzelle ebenfalls eine periodische Funktion des Anstellwinkels φ dieser Geraden. Die Periode ist in allen Fällen gleich π.
Treffen diese vier Feuerraten im visuellen Cortex in den Ecken eines Vierecks ein, und empfängt ein beliebiges Neuron innerhalb dieses Vierecks die neuronale Erregung, so ist die resultierende Feuerrate bei additiver Überlagerung der Einzelfeuerraten ebenfalls eine periodische Funktion des Anstellwinkels φ mit der Periode π.
Jede periodische Funktion hat Extremwerte, sowohl Minima als auch Maxima.
Dies gilt auch, wenn es eine abstandsabhängige Dämpfung gibt, weil die Dämpfung jedes Erregungsbeitrages bei unveränderlichem Ort aller beteiligten Neuronen eine konstante Größe ist.
Erfolgt die abstandsabhängige Dämpfung durch eine konkave Funktion, so existiert ein globales Maximum, dessen Lage ebenfalls eine periodische Funktion des Anstiegswinkels φ der Geraden ist. Für genau einen Anstellwinkel φmax erreicht die Erregung des Neurons ein globales Maximum.
Somit ist das Verhalten der Orientierungssäulen im primären visuellen Cortex eine Gesetzmäßigkeit. Die Erregung eines (ortsfesten) Neurons ist eine periodische Funktion des Anstellwinkels.
Nach diesen Darlegungen wäre es eigentlich nicht mehr nötig, die langwierigen Berechnungen durchzuführen, die zeigen, dass Orientierungssäulen genau das Verhalten haben, welches man messtechnisch nachgewiesen hat. Jeder Mathematiker müsste an dieser Stelle bereits zustimmen, dass das beobachtete Verhalten gesetzmäßig auftreten muss. Das beobachtete Verhalten der Neuronen in Orientierungssäulen ist auch nur zu erklären, wenn die Übertragungsfunktion für neuronale Erregungen eine konkave Funktion des Abstandes ist.
Nach der Darlegung der elementaren mathematischen Grundlagen für Divergenzmodule und die in ihnen stattfindende Maximalcodierung von Parametern können wir konkrete Divergenzmodule des menschlichen Gehirns konkreter analysieren.
Divergenzmodule mit vertikaler Signalmischung
4.1.2. Das Farbmodul mit vertikaler Signalmischung
Die folgende Abbildung soll den Aufbau eines Divergenzmoduls mit vertikaler Signalüberlagerung im primären visuellen Cortex zeigen. Wir wählen als Beispiel nur das Farbsystem Rot-Grün aus. Die Schichtdicke S4 der Outputschicht S4-Grün hat bereits zugenommen, ihre Outputneuronen sind signalkompatibel sowohl mit den Inputsignalen der Grün-Rezeptoren als auch mit denen der Rot-Rezeptoren. Daher empfängt jedes Outputneuron sowohl Input von unten als auch von oben.
Jedem Bildpunkt der Retina ist so ein Farbmodul zugeordnet, wenn man davon ausgeht, dass die visuelle Cortexrinde lediglich ein Dickenwachstum entwickelte. Später wird der Fall untersucht, dass ein Breitenwachstum stattfand, und zuletzt untersuchen wir den Fall, dass es sowohl ein Dickenwachstum als auch ein Breitenwachstum gab.
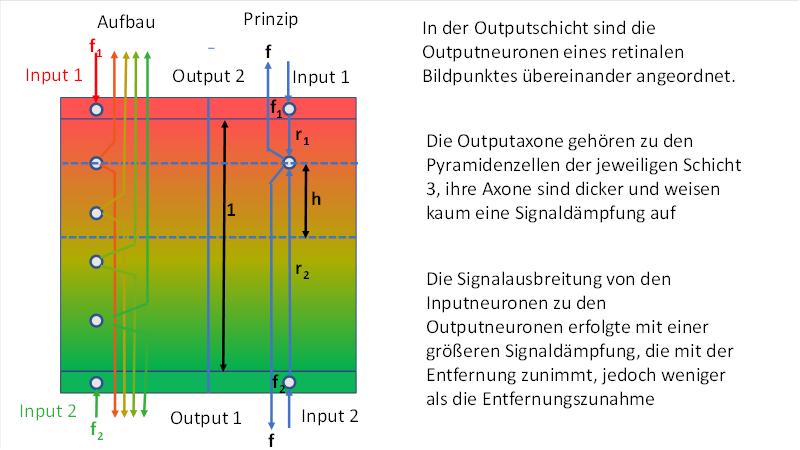
Abbildung 9: Divergenzmodul für Farbsehen Rot-Grün
Die obige Abbildung ist zweigeteilt durch eine blaue Linie: Links ist ihr wirklicher Aufbau im Cortex dargestellt, rechts das Prinzip der Signalausbreitung und die Größen, die für die mathematische Beschreibung erforderlich sind.
Symbolisch dargestellt sind die zu einem Bildpunkt der Retina gehörenden zwei Outputneuronen mit zueinander inversen Signalen f1 = Rot-On/Grün-Off und f2 = Grün-On/Rot-Off nach der gängigen Gegenfarbentheorie. Wir verkürzen die Schreibweise und sprechen nachfolgend vereinfachend vom Grün-On-Neuron und vom Rot-On-Neuron.
Genau auf der Verbindungslinie der zwei Inputneuronen sind in etwa gleichem Abstand die Outputneuronen verteilt.
Input- und Outputneuronen bilden so eine schmale Augendominanzsäule.
Es sei f1 die Feuerrate des Rot-On-Neurons und f2 die Feuerrate des Grün-On-Neurons.
Ursprünglich hatte jede Inputschicht der Klasse 4 eine ihr zugeordnete Outputschicht der Klasse 3 genau darüber - es teilten sich also sowohl die Submodalitäten als auch die Schichten der Neuronenklasse und ordneten sich für jede Submodalität neu.
So entstanden die Inputschichten 3-Grün-On und 3-Rot-On.
Da aber der Rot-On-Rezeptor ein Abkömmling des Grün-On-Rezeptors war, waren sie beide signalkompatibel, also signalverwandt. Daher konnten die Outputneuronen der unteren Outputschicht S3-Grün-On auch die Erregung von den Outputneuronen der oberen Schicht S4-Rot-On empfangen. Die Outputschicht S3-Rot-On wurde überflüssig und bilde sich im Verlauf der Evolution zurück.
Die von den Inputneuronen in das System übertragene Erregung möge sich in der Schicht ausbreiten und jedes Neuron der Augendominanzsäule erreichen. Hierbei gelte die spezifizierte Kabelgleichung für marklose Axone.
Wir betrachten ein ausgewähltes Outputneuron, welches von den Inputneuronen den Abstand r1 und r2 haben möge. Die Erregungen von beiden Inputneuronen erregt das Outputneuron entsprechend der Formel
![]()
Hierbei gilt:
![]()
![]()
Wie bereits an der Hesse-Matrix der Übertragungsfunktion eines einzelnen Neurons gezeigt, existiert ein globales Maximum der Feuerrate. Wir möchten wissen, wo sich das Outputneuron mit der maximalen Erregung bei vorgegebenen Feuerraten f1 und f2 befindet.
Das Maximum ermitteln wir durch Nullsetzen der Ableitung.
Wir beachten, dass der Abstand zwischen den zwei Inputneuronen gleich 1 ist, das Outputneuron hat den Abstand h von der Mitte der Outputschicht.
Dann gilt:
![]()
Wir bilden die Ableitung nach der Variablen h und setzen sie gleich Null.
![]()
Zunächst setzen wir die Größen r1 und r2 wieder ein und erhalten ein Zwischenergebnis für den späteren Gebrauch im Konvergenzgitter.
![]()
![]()
![]()
Gleiche Faktoren auf beiden Seiten fallen weg.
![]()
![]()
![]() (Hauptformel
1)
(Hauptformel
1)
Wir setzen wieder die Gleichungen für r1 und r2 ein und erhalten
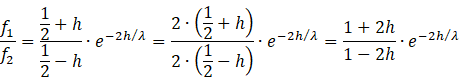
![]() (Hauptformel 2)
(Hauptformel 2)
Wir logarithmieren
![]()
![]()
Wegen
![]()
Für alle Werte x mit |x| < 1.
Weil h im Intervall von - ½ bis + ½ liegt, als 2h im Intervall von -1 bis +1, können wir diese Formel anwenden und erhalten die Beziehung
![]()
Dies ist gleichbedeutend mit der Gleichung
![]() (Hauptformel
3)
(Hauptformel
3)
Die Hauptformeln 1 und 2 erlauben es, das notwendige Feuerratenverhältnis für ein Outputneuron im Farbmodul zu berechnen, welches auf der Höhe h die maximale Erregung innerhalb der gesamten Farbsäule einnimmt.
Die Funktion
![]()
erlaubt es, das Feuerratenverhältnis in Abhängigkeit von der Höhe x zu untersuchen. Wir bedenken, dass x eine positive Zahl zwischen 0 und ½ sein muss, denn die Gesamtdicke des Farbmoduls haben wir gleich 1 gesetzt. Die Funktion f besitzt beispielsweise für den Wert λ = 5 folgendes Aussehen:
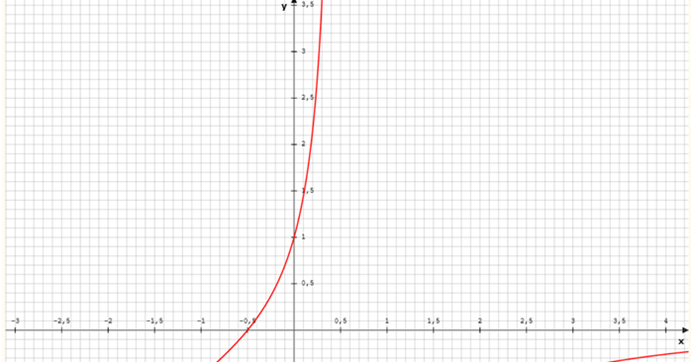
Abbildung 10: Feuerratenverhältnis im Divergenzgitter als Funktion von x
Die Größe x im Diagramm entspricht der Höhe h im Divergenzmodul und kann Werte zwischen -0,5 und +0,5 annehmen.
Für x = 0 besitzt die Funktion den Funktionswert 1, dann sind beide Feuerraten gleich groß und das Maximalneuron befindet sich in der Mitte zwischen beiden Inputneuronen.
Für h = - ½ ist der Funktionswert gleich null. In diesem Fall ist f1 = 0. Für h= + ½ wird der Funktionswert unendlich. Hier wäre f2 = 0.
Zu beachten ist in dem obigen Diagramm, dass nur positive Funktionswerte auftreten können. Beide Feuerraten sind positiv, daher ist ihr Quotient ebenfalls positiv.
Das beschriebene Divergenzmodul mit vertikaler Signalmischung und dem Input der rot- und grünempfindlichen Sehrezeptoren erzeugt einen maximumcodierten Output in der Augendominanzsäule. Das maximal erregte Outputneuron verschlüsselt das Mischungsverhältnis der Farben Rot und Grün, für die beide Sehrezeptoren unterschiedliche Empfindlichkeitskurven besitzen. Ändert sich das Mischungsverhältnis, so wandert das Erregungsmaximum nach oben bzw. unten. Damit erkennt das Divergenzmodul im Wellenlängenbereich Rot-Grün eine Farbe, also auch eine Wellenlänge.
Drei solcher Module befinden sich im Cortex übereinander, jedes deckt einen anderen Wellenlängenbereich ab. Wir wollen die Gesamtheit aus diesen drei Modulen als Farbmodul bezeichnen. Dann gilt folgende Aussage:
Das dreiteilige Farbmodul mit vertikaler Signalausbreitung im visuellen Cortex kann Farben im Wellenlängenbereich Rot-Grün-Blau erkennen und durch einen maximumcodierten Output bezüglich der Wellenlänge charakterisieren. Es kann Farben erkennen.
Wenn der Output dieses Farbmodul (nach oben oder nach unten) verlässt und es anschließend eine laterale Nachbarhemmung zwischen den Outputsignalen gibt, bleiben nur noch diejenigen Neuronen aktiv, die sich in unmittelbarer Nähe des Erregungsmaximums befinden. Die Erregung der übrigen Neuronen in der gesamten Augendominanzsäule fallt der lateralen Hemmung zum Opfer. So entsteht ein Outputvektor mit spärlicher Codierung.
Der Output dieses Moduls kann einer neuen Modalität zugeordnet werden, die es im Gehirn vorher so nicht gab: der Farbe.
Nun begann ein Prozess, der letztlich beim Homo sapiens einen vorläufigen Höhepunkt erreichte. Die neue Modalität Farbe verselbständigte sich. Wie übrigens alle weiteren, neuen Modalitäten, die in den verschiedenen cortikalen Divergenzgittern entstanden.
Wie haben wir uns die Verselbständigung vorzustellen?
Etwa so, als wäre eine neue Rezeptorenart entstanden, die diese neue Modalität hervorbrachte. Die Outputneuronen des Divergenzgitters übernahmen die Rolle von Rezeptoren. Ihr Output zog kopfwärts nach oben und begründete ein neues Leitersegment im alten Strickleitersystem. Über dem alten, cortikalen Segment, welches eigentlich das letzte Segment im alten Strickleitersystem war, entstanden neue Leitersegmente für die neuen Modalitäten. Doch da der Platz nach oben durch den Schädelknochen begrenzt war, bogen die neu entstehenden Leiterelemente der neu entstandenen Modalitäten einfach seitlich ab. So wuchs der Cortex auch seitlich, die Lobi erfuhren eine starke seitliche Ausdehnung und bogen sich dabei, den Schädelknochen folgend, zu Halbschalen. Platzmangel führte dann auch hier zur Faltenbildung. Beschrieben wird diese Entwicklung in einem eigenen Kapitel am Ende dieses Buches ausführlicher. So entstand neben dem primären Cortex der sekundäre Cortex, der auch als Assoziationscortex bezeichnet wird und ein Zeichen von höherer Intelligenz ist.
Das Divergenzmodul mit vertikaler Signalüberlagerung war jedoch ausbaufähig. Im Verlauf der Evolution wurde das Dickenwachstum der Outputschichten durch ein Wachstum in die Breite ergänzt. Spezies, denen dies gelang, konnten Divergenzmodule hervorbringen, in denen sich die Erregung sowohl vertikal als auch horizontal ausbreitete. Eine gesetzmäßige Folge dieser seitlichen Signaldivergenz war im motorischen Cortex die Herausbildung der Gelenkwinkelanalyse durch maximalerregte Neuronenpopulationen, die bei periodischer Winkeländerung um ein Zentrum rotierten. Im visuellen Cortex entstanden Orientierungssäulen, deren Maximalerregung die Neigung einer Geraden im Gesichtsfeld repräsentierte. Hierbei wurde der Anstiegswinkel der Geraden von der magnocellularen Outputschicht hinsichtlich ihrer Helligkeit (Weiß, Graustufen, Schwarz) analysiert, während die parvocellularen Schichten die Farbe der geneigten Geraden erkennen konnten. Dies wird im Kapitel über die Divergenzmodule mit räumlicher Signalüberlagerung erklärt und bewiesen. Doch zuvor betrachten wir das Helligkeitsmodul mit vertikaler Signalüberlagerung.
Es erscheint mit notwendig, die durch das Farbmodul mit vertikaler Signalmischung entstehenden Outputsignale einer neuen Modalität zuzuordnen: der Farbe.
Zunächst nur Mischfarben Rot-Grün, später beim Menschen der Spektralbereich von Rot über Gelb und Grün bis zum Blau gab es nun Neuronen, deren maximale Aktivität einer bestimmten Farbe zugeordnet war.
Obwohl in dieser Monografie das Farbmodul mit vertikaler Signalmischung zuerst vorgestellt wurde, gab es ein Modul mit vertikaler Signalmischung, welches viel früher entstanden sein muss. Denn das Farbmodul benötigt zwei unterschiedliche Arten von Sehfarbstoffen. Zunächst gab es nur einen, und der ermöglichte das Hell/Dunkel-Sehen. Das Wirkprinzip ist das Gleiche wie beim Farbmodul.
4.1.3. Das Helligkeitsmodul mit vertikaler Signalmischung
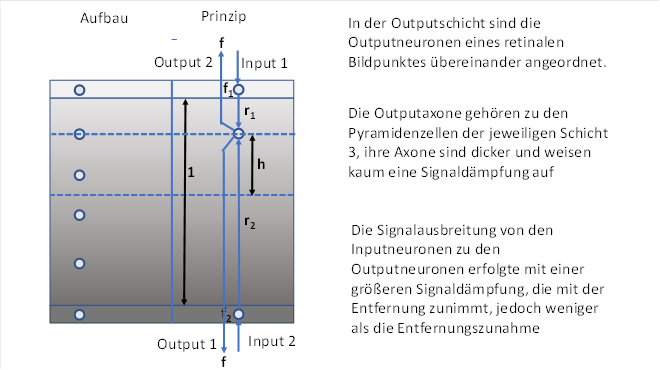
Abbildung 11: Helligkeitsmodul
Symbolisch dargestellt sind die zu einem Bildpunkt der Retina gehörenden zwei Outputneuronen mit zueinander inversen Signalen. Genau auf der Verbindungslinie der zwei Inputneuronen sind in etwa gleichem Abstand die Outputneuronen verteilt.
Input- und Outputneuronen bilden so eine schmale Augendominanzsäule.
Es sei f1 die Feuerrate des Hell-On-Neurons und f2 die Feuerrate des Dunkel-On-Neurons.
Wir gehen von einer Helligkeit u aus und ordnen f1 und f2 folgende Werte zu. f1 sei die Helligkeit der Hell-On-Ganglienzelle, f2 die der Dunkel-On-Ganglienzelle. Diese Ganglienzelle bezeichnen wir auch als Hell-Off-Zelle. Wir gehen hier davon aus, dass es die Submodalität Dunkel-On zuerst gab. Daher bildet sie im Thalamus und Cortex die untere der beiden Inputschichten.
Ursprünglich hatte jede Inputschicht der Klasse 4 eine ihr zugeordnete Outputschicht der Klasse 3 genau darüber - es teilten sich also sowohl die Submodalitäten als auch die Schichten der Neuronenklasse und ordneten sich für jede Submodalität neu.
So entstanden die Inputschichten 4-Hell-On und 4-Hell-Off sowie die Schichten zwei Outputschichten 3-Hell-On und 3-Hell-Off.
Da aber der Hell-On-Rezeptor ein Abkömmling des Dunkel-On-Rezeptors war, waren sie beide signalkompatibel, also signalverwandt. Daher konnten die Outputneuronen der unteren Outputschicht S3-Dunkel-On auch die Erregung von den Outputneuronen der oberen Schicht S4-Hell-On empfangen. Die Inputschicht S4-Hell-On wurde überflüssig und bilde sich im Verlauf der Evolution zurück.
Die von den Inputneuronen in das System übertragene Erregung möge sich in der Schicht ausbreiten und jedes Neuron der Augendominanzsäule erreichen. Hierbei gelte die spezifizierte Kabelgleichung für marklose Axone.
Wir betrachten ein ausgewähltes Outputneuron, welches von den Inputneuronen den Abstand r1 und r2 besitzt. Die Erregungen von beiden Inputneuronen erregen das Outputneuron entsprechend der Formel
![]()
Wie bereits an der Hesse-Matrix der Übertragungsfunktion gezeigt, existiert ein globales Maximum der Feuerrate. Wir möchten wissen, wo sich wir bei vorgegebenen Feuerraten f1 und f2 das Outputneuron mit der stärksten Erregung befindet.
Das Maximum ermitteln wir durch Nullsetzen der Ableitung.
Wir beachten, dass der Abstand zwischen den zwei Inputneuronen gleich 1 ist, das Outputneuron hat den Anstand h von der Mitte der Outputschicht.
Dann gilt:
![]()
Dies ist genau die Gleichung, die wir bereits beim Farbmodul mit vertikaler Signalmischung erhalten haben. Aus ihr werden sich daher die gleichen Resultate ableiten lassen.
![]() (Hauptformel
1)
(Hauptformel
1)
![]() (Hauptformel
2)
(Hauptformel
2)
![]() (Hauptformel
3)
(Hauptformel
3)
Diese Formel erlaubt es uns, aus den Feuerraten f1 und f2 zu berechnen, an welcher Stelle h das Outputneuron unter allen anderen die maximale Feuerrate hat. Sein Output codiert damit den Feuerratenquotienten. Anstelle der Farben kann dieses Modul die Helligkeit bewerten. Der Formalapparat ist der gleiche. Ein Modul kann also beliebige Modalitäten bewerten, solange sie in zwei Submodalitäten zerfallen, von denen eine vom On-Typ und die andere vom Off-Typ ist.
Ein solches Modul befindet sich im Cortex genau unter dem dreigeteilten Farbmodul. Wir werden es als Helligkeitsmodul mit vertikaler Signalmischung bezeichnen.
Dann gilt folgende Aussage:
Das Helligkeitsmodul mit vertikaler Signalausbreitung im visuellen Cortex kann Helligkeiten als Grautöne im Intervall von Weiß bis Schwarz erkennen durch einen maximumcodierten Output bezüglich der Wellenlänge charakterisieren. Es kann Helligkeiten erkennen.
Wenn der Output dieses Helligkeitsmoduls (nach oben oder nach unten) verlässt und es anschließend eine laterale Nachbarhemmung zwischen den Outputsignalen gibt, entsteht ein Outputvektor mit spärlicher Codierung. Es bleiben nur noch diejenigen Neuronen aktiv, die sich in unmittelbarer Nähe des Erregungsmaximums befinden. Die Erregung der übrigen Neuronen in der gesamten Augendominanzsäule fallt der lateralen Hemmung zum Opfer.
4.1.4.Neue Modalitäten im vertikalen Divergenzmodul
Durch die Divergenzmodule mit vertikaler Signalmischung, kurz vertikale Divergenzmodule, entstanden neue Outputarten. Mit jedem neuen Neuron, welches in der Outputschicht entstehen konnte, wurde die neue Outputart durch eine neue Klasse von Neuronen erweitert, die etwas konnten, was vorher nicht ging: Farben erkennen, Helligkeitsstufen erkennen, Signalmischungen erkennen. Dies erwies sich als so bedeutsam und vorteilhaft, dass sich die jeweils neue Neuronenklasse zu einer eigenen Modalität verselbständigte. So entstand die Modalität Farbe, Helligkeit, aber auch Duft oder Geschmack. Das wichtigste Erkennungsmerkmal einer neuen Modalität ist ihre Aufspaltung im neuronalen System.
Die Verselbständigung einer neuen Modalität - das Erringen der Modalitäteneigenschaft - hatte weitreichende Konsequenzen. Gleichartige Modalitäten (und Submodalitäten wie hier die Farbe) ordnen sich neu. Dies bedeutet, dass die verschiedenen Farben sich im Gehirn zu trennen begannen. Wenn eine Augendominanzsäule für die Farbwahrnehmung eines Retinapixels beispielsweise aus 20 Outputneuronen bestand, die wie eine Perlenschnur von oben nach unten durch alle drei parvocellularen Cortexschichten der Klasse 3 verlief, so teilte sich der Output dieser 20 Neuronen auf 20 neue sekundäre Gehirngebiete auf. Es gab dann 20 Retinaabbildungen, aber jede von ihnen war nur für eine der erkennbaren Farben zuständig.
War also links ein grüner Apfel zu sehen und rechts eine rote Erdbeere, so kam das Apfelbild in der Retinaabbildung an, die auf Grün reagierte, während das rote Erdbeerbild in einer Retinaabbildung erschien, die auf Rot reagierte und sich an einer völlig anderen Stelle des Cortex befand. Es ist etwa so, als würde man einen Farbfilm gleichzeitig auf 20 verschiedenen Fernsehgeräten betrachten, von denen jedes Gerät nur einen Spektralbereich anzeigt, der ein Zwanzigstel des sichtbaren Farbspektrums umfasst. Deshalb sind wir berechtigt, den Output dieses Moduls als Farbmodalität zu bezeichnen. Der Begriff Farbmodalität bezeichnet die Zugehörigkeit von Neuronen zu diesem Farbmodul, aber ebenso die Outputsignale dieses Moduls.
Es wird auffallen, dass der mathematische Apparat beim Farbsehen und beim Helligkeitssehen völlig identisch ist. Auch jede andere Modalität kann die Signale ihrer Rezeptoren in solche Divergenzgitter einspeisen. Sie sollten jedoch immer als inverse Signalklassen vorliegen. Dann kann die Stärke der Urgröße aus der Lage des maximal erregten Neurons abgeleitet werden und die Modalität ist einer Feinanalyse zugänglich.
Deshalb gibt es in vielen Rezeptorsystemen On-Signale und Off-Signale. Und da, wo sie bei einer Modalität fehlten, konnte das Spinocerebellum und das Vestibulocerebellum eine Signalinversion durchführen. So gab es zu jedem motorischen Signal im Cortex ein inverses Signal im Cerebellum, welches auch den Cortex erreichte. So konnte beispielsweise die Muskelkraft analysiert werden.
Damit ein motorisches Signal und das aus ihm im Cerebellum invertierte Signal jedoch in der gleichen Cortexhälfte ankamen, musste das motorische Signal vor dem Erreichen des Cerebellums, also wahrscheinlich noch im Rückenmark, die Körperseite wechseln, bevor es zum Cortex aufstieg. Es musste also (für motorische Signale) eine Signalkreuzung auf Rückenmarksebene geben, die ausschließlich den Cortex versorgte, während die ungekreuzten Signale den Olivenkern erreichten, so ebenfalls zur anderen Seite zogen und im Cerebellum invertiert wurden. Die aufsteigende Projektion vereinigte die Originalsignale und die aus ihnen durch Inversion im Cerebellum gewonnenen Signale in einem gemeinsamen motorischen Divergenzmodul mit vertikaler Signalüberlagerung. So konnte etwa die Muskelkraft fein dosiert werden, weil sie messbar wurde.
In den Outputschichten von Divergenzmodulen findet eine Extremwertcodierung statt. Das maximal erregte Neuron bewertet die jeweilige Submodalität hinsichtlich ihrer Signalstärke. Seine Aktivität ruft das hervor, was Neurologen und Philosophen als Qualia hinsichtlich eines elementaren Reizes bezeichnen.
Damit offenbart sich ein grundlegendes Verständnisproblem. Helligkeit ist für Physiker und viele Naturwissenschaftler der Energiegehalt des wahrgenommenen Lichtes. Farbe ist für Physiker und für Naturwissenschaftler die Wellenlänge von Licht. Geruch ist das Intensitätsverhältnisses von Stoffen, die durch Verdunstung in der Luft vorhanden sind.
Neurologen, Hirnforscher und Philosophen diskutieren auch über Helligkeit, Farbe oder Geruch. Sie meinen jedoch nicht die außerhalb unseres Gehirns vorhandenen Zustände von Lichtenergie, Lichtwellenlänge oder Mischungsverhältnis von Duftstoffen in der Luft. Sie meinen den geistigen Zustand im Gehirn, den sie als Qualia bezeichnen und der sich einer wissenschaftlichen Erklärung zu entziehen scheint.
In dieser Monografie wird gezeigt, dass die Helligkeit, die Farbe, der Geruch, die Muskelspannung und vieles andere in einem vertikalen Divergenzgitter zur spezifischen Maximalerregung eines Neurons führt, welches sich in einer mathematisch definierten Höhe in der Outputschicht der jeweiligen Submodalität befindet.
Bei denjenigen Menschen, deren Outputschicht bezüglich einer Submodalität die gleiche Dicke und die gleiche Neuronenanzahl aufweist sowie die gleiche topologische Anordnung, werden genau die gleichen Neuronen aktiviert, wenn eine bestimmte Submodalität, z. B. die Lichtfarbe, analysiert wird. Diese Gehirnabschnitte oder Gehirnschichten könnten dann bei verschiedenen Menschen (theoretisch) ausgetauscht werden, ohne die Analyseergebnisse zu verfälschen.
Insofern sind die primären Hirnrinden des menschlichen Cortex bei verschiedenen Menschen gleichartig aufgebaut und im Prinzip kompatibel zueinander. Hat jemand jedoch eine Outputschicht S3 in einem primären Cortexgebiet, die deutlich dicker ist als bei den meisten Menschen, so kann er die betreffende Modalität (Farbe, Helligkeit, Tonhöhe, -) etwas feiner analysieren, erhält bei der Analyse also mehr Zwischenstufen.
Fehlt eine Outputschicht S3, so ist der betreffende -blind- (unempfindlich) für die zugehörige Submodalität (Farbenblindheit, Blindheit, Geruchsstörung, Bewegungsstörung, -), aber auch ein Ausfall von Rezeptoren kann dies bewirken. Besonders tragisch ist es, wenn dies durch angeborene Gendefekte verursacht wird, weil die Betroffenen dann zeit des Lebens diese Wahrnehmung niemals erleben. Die fehlende Erfahrung macht es besonders schwer, den Betroffenen die Existenz der für sie fehlenden Submodalität zu erklären.
Die Helligkeits- und Farbmodule mit vertikaler Signalüberlagerung erweiterten ihren Analyseumfang deutlich, als ein Wachstum der Outputschichten in horizontaler Richtung erfolgte. Zu erkennen war die starke seitliche Ausdehnung der Cortexschichten an der Größenzunahme des Cortex und an der eintretenden Faltenbildung. Legt man eine Schicht in Falten, so kann sie auf gleichem Raum mehr Fläche besitzen. Anhand der Ausprägung der Faltenbildung der Cortexschicht kann darauf geschlossen werden, ob das betreffende Tier evolutionär älter ist (weniger Falten, kleine Größe) oder evolutionär jünger (mehr Falten, größere Fläche).
Das Prinzip der Divergenzgitter mit räumlicher Signalüberlagerung wird zunächst am Beispiel der Motorik beschrieben. Alle Rezeptoren, die die Motorik kontrollieren, enden in verschiedenen sensorischen Cortexschichten. Diese bilden genau die Divergenzmodule, die nachfolgend beschrieben werden.
Im nachfolgenden Abschnitt wird untersucht, welche Folgen das seitliche Schichtenwachstum hatte, wobei so getan wird, als hätte es ein Dickenwachstum nicht gegeben. Dies ist zwar realitätsfremd, aber es ist oft nützlich, zwei verschiedene Prozesse entkoppelt und für sich allein zu betrachten. Daher analysieren wir nun das Divergenzgitter mit seitlicher Signalausbreitung und tun so, als ob es eine vertikale Signalausbreitung nicht gäbe, weil die Outputschicht eine einlagige Neuronenschicht sei.
Erst danach werden das vertikale und das seitliche Schichtenwachstum im Cortex analysiert und das Prinzip von cortikalen Divergenzmodulen mit räumlicher Signalausbreitung behandelt. Durch diese Vorgehensweise hoffe ich, diejenigen Leser nicht zu überfordern, die mit dem mathematischen Formelapparat schon genügend Probleme haben werden.
Monografie von Dr. rer. nat. Andreas Heinrich Malczan