Gehirntheorie des Menschen
für das Human Brain Projekt der Europäischen Union
ISBN 978-3-00-068559-0
4.2 Divergenzmodule mit seitlicher Signalausbreitung
Das motorische Divergenzmodul mit seitlicher Signalausbreitung
In motorischen Cortexfeldern beobachtet man Erregungsmaxima, die umherwandern, wenn körperliche Bewegungen stattfinden.
Im Fachbuch "Neurowissenschaften" [1] von Eric R. Kandel, James H. Schwartz und Thomas M. Jessell, Spektrum Akademischer Verlag 1995, werden im Kapitel 29 die Forschungsergebnisse zur Willkürmotorik dargestellt. Es wird gezeigt, dass Neuronen im primären motorischen Cortex die Kraft und die Richtung von Willkürbewegungen kodieren.
In einer Versuchsanordnung bewegten Affen einen Stab mit dem Arm zu einem von mehreren Zielpunkten, die in unterschiedlichen Richtungen um eine zentrale Startposition angeordnet waren.
Während der Bewegungen wurden die Feuerraten von sehr vielen cortikalen Neuronen in dem Gebiet gemessen, in dem die Signale der Rezeptoren der zugehörigen Muskeln und Gelenke eintrafen.
Es zeigte sich, dass die Bewegungsrichtung nicht von einzelnen Neuronen, sondern von Neuronenpopulationen codiert wird. Diese Neuronenpopulationen sind etwa kreisförmig um ein Zentrum angeordnet. Je nach Bewegungsrichtung ist jedoch immer nur eine Neuronenpopulation aktiv. Deren Lage zum Zentrum codiert die Bewegungsrichtung derart, dass der Populationsvektor weitgehend mit der Bewegungsrichtung übereinstimmt.
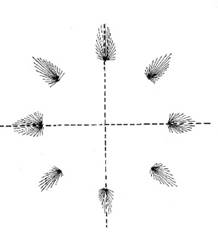
Abbildung 12: Codierung der Bewegungsrichtung durch aktive Neuronenpopulationen
Abb. 12 nach [1], Seite 545, Abbildung 29.3
Ist es möglich, dieses Phänomen wissenschaftlich zu erklären? Kann man aus der Position eines solchen Erregungsmaximums möglicherweise auf die Gelenkwinkel schließen, die mit der Bewegung verbunden sind. Kann man vielleicht diese Gelenkwinkel aus der ermittelten Erregungsposition errechnen, also einen direkten mathematischen Zusammenhang zwischen Erregungsmaximum und Gelenkwinkel herstellen?
Ja, dies ist prinzipiell möglich. Das motorische Divergenzmodul schafft den informationstheoretischen Zugang zu dieser Thematik.
Wir betrachten beispielhaft das Schultergelenk, welches es uns ermöglicht, den Arm nach vorn zu heben und zu senken, aber ebenso ihn seitlich zu hin- und her zu bewegen. Dazu sind vereinfacht vier Muskeln nötig, von denen jeweils zwei ein Muskelpaar bilden, welches antagonistisch gegeneinander arbeitet. Die Muskeln werden von Motoneuronen angesteuert, deren Feuerrate über den Grad der Kontraktion entscheidet.
Dem ersten Muskelpaar ordnen wir die Feuerraten f1 und f3 zu. Diese Muskeln wirken gegeneinander, der eingestellte Gelenkwinkel der ersten Ebene hängt vom Feuerratenverhältnis ab. Für diese zwei Rezeptorsignale wählen wir den exponentiellen Ansatz gemäß
Dies ist immer möglich, wenn man folgenden Ansatz wählt:
![]()
![]()
Es gilt
Die Größe fm interpretieren wir als mittlere Feuerrate, sie genügt der Gleichung
![]()
Die Größe u bezeichnen wir als Urgröße des Gelenks. Sie repräsentiert (beispielsweise) den Gelenkwinkel, mit dem wir den Arm nach vorn heben bzw. senken. Der Wert u = 0 möge erreicht sein, wenn der Arm gerade nach vorn ausgestreckt wird. Positive Werte von u führen zum Anheben des Armes nach vorn, negative zum Absenken. Für das Heben des Arms ist f1 zuständig, für das Absenken dagegen f3.
Dem zweiten Muskelpaar ordnen wir die Feuerraten f2 und f4 zu. Auch hier wählen wir den exponentiellen Ansatz
![]()
![]()
Hier ist die Urgröße v für das seitliche Bewegung des Arms zuständig, die Feuerrate f2 bewege den Arm nach links, die Feuerrate f4 nach rechts. Im Allgemeinen werden sich die Bewegungen aus ihren zwei Komponenten zusammensetzen, denn das Gelenk hat zwei Freiheitsgrade.
Den zwei Urgrößen u und v können wir im kartesischen Koordinatensystem darstellen.
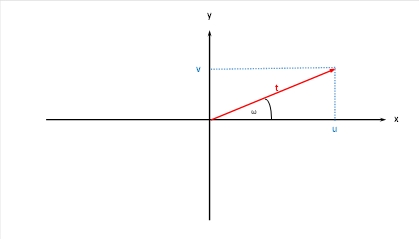
Abbildung 13: Urgrößen u und v im kartesischen Kooedinatensystem
![]()
![]()
Wir bezeichnen den Winkel ω als Phasenwinkel und t als Radiusvektor im Urgrößendiagramm.
Die vier Feuerraten f1 bis f4 erreichen über die Axone der erzeugenden Rezeptoren im Neuralrohr je vier Neuronen der Klasse 3 und 4. Die Neuronen der Klasse 4 projizieren kopfwärts, wobei die Feuerrate unverändert bleiben möge, so dass unter Zwischenschaltung weiterer Neuronen der Klasse 4 der (sensorische) Cortex erreicht wird. Dort werden vier Neuronen der Klasse 4 mit diesen vier Feuerraten f1 bis f4 erregt. Entlang der gesamten Projektionsstrecke mögen die Feuerraten f1 bis f4 unverändert bleiben.
Wir müssen nun noch entscheiden, welche Neuronenschicht den Input empfängt, wo sich die Interneuronen befinden müssen und welche Schicht die Outputschicht ist. Wir gehen davon aus, dass die Schicht vier die Inputschicht ist. In dieser Schicht befinden sich auch alle Interneuronen, die an der Signalausbreitung in der Schicht vier beteiligt sind.
Wir betrachten in diesem Unterkapitel den Sonderfall, dass die Outputschicht 3 verwendet wird. Wir gehen - was die Evolution betrifft - von dem sehr frühen Fall aus, dass die Outputschicht eine sehr dünne Schicht ist. Die Inputneuronen der Nachbarschicht 4 mögen größere Abstände zueinander besitzen. Dies bedeutet, dass sich diese Schicht - und mit ihr alle anderen Schichten - flächenmäßig ausdehnen. Dadurch wächst der Gehirnumfang der zugehörigen Cortexareale.
Später werden wir in einem folgenden Kapitel diese Beschränkung aufheben und analysieren, welche Effekte ein Dickenwachstum der Outputschicht 3 hat.
Als Outputschichten kommen zwei in Betracht - die Schicht 3 projiziert den Output zum motorischen Zentrum (zum motorischen Cortex), die Schicht 6 projiziert zu den Mittelwertkernen und die Schicht 2 zur kontralateralen Seite. Wir unterstellen, dass die Schicht 3 in diesem Modul die eigentliche Arbeit macht. Ihr Output wird über Pyramidenzellen der Schichten 2 bzw. 6 zu den nachfolgenden Auswerteeinheiten weitergeleitet.
In einem linearen Divergenzgitter überlagerten sich die Erregungen von zwei Neuronen. In einem ebenen Divergenzgitter erfolgt die Ausbreitung der Erregung in der Fläche, in der sowohl die Inputneuronen als auch die Outputneuronen verteilt sind.
Wir vergleichen beide Divergenzgitter in nachfolgender Abbildung.
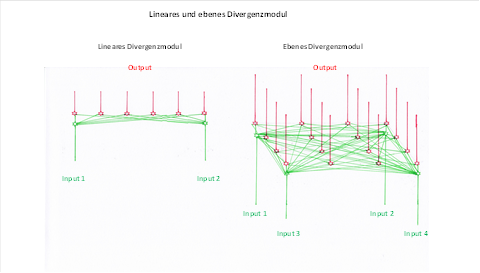
Abbildung 14: Lineares und ebenes Divergenzmodul im Vergleich
Das ebene Divergenzgitter möge vier Inputneuronen besitzen. Wir ordnen die vier Inputneuronen N1 bis N4 in einem Koordinatensystem so an, wie es die nachfolgende Abbildung zeigt.
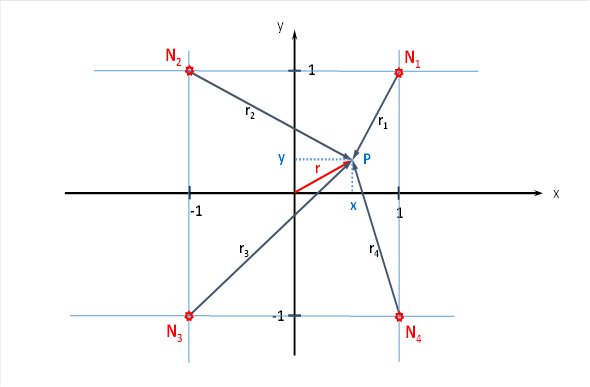
Abbildung 15: Ebenes Divergenzgitter mit vier Inputneuronen
Der Radiusvektor r sowie der Phasenwinkel α und die Größen x und y sind durch folgende Gleichungen verknüpft.
![]()
![]()
![]() .
.
Hierbei sei α der Winkel, den der Radiusvektor r mit der x-Achse bildet.
Im Punkt P mit den Koordinaten P = (x; y), zu dem der Radiusvektor r gehört, möge sich nun genau dasjenige Neuron befinden, welches unter allen Neuronen im gegebenen Quadrat die größte Erregung aufweist. Damit dies passiert, muss die Gelenkstellung eine ganz bestimmte Position einnehmen. Wir versuchen nun, aus den Koordinaten x und y dieses globalen Maximums die Werte u und v der beiden Urgrößen zu bestimmen.
Wir setzen voraus, dass sich die neuronale Erregung von den vier Inputneuronen in alle Richtungen ausbreitet und im Punkt P(x; y) mit den Koordinaten x und y ihr absolutes Maximum annimmt. Dass es ein solches gibt, ist theoretisch bereits bewiesen (siehe Bemerkungen zur Hessematrix).
Aus den zu erfüllenden Extremwertbedingungen ermitteln wir die Werte der nötigen Urgrößen u und v.
Für die beteiligten Radiusvektoren r1 bis r4 erhält man über den Satz des Pythagoras nachfolgende Gleichungen:
![]()
![]()
![]()
Wir berücksichtigen die nachfolgenden Beziehungen
![]() .
.
Nun ersetzen wir in den obigen Gleichungen die Summe und die Differenz von x und y wie folgt
![]()
![]()
Und erhalten so
![]()
![]()
![]()
![]()
Befinden sich
die vier Inputneuronen in den Punkten ![]() ,
,
![]() ,
,
![]() und
und
![]() und
erhalten die Erregungen mit den Feuerraten f1, f2, f3
und f4, so kann unter unseren Voraussetzungen die Feuerrate f des
Outputneurons im Punkt
und
erhalten die Erregungen mit den Feuerraten f1, f2, f3
und f4, so kann unter unseren Voraussetzungen die Feuerrate f des
Outputneurons im Punkt ![]() als
Summe der vier beteiligten Erregungen berechnet werden:
als
Summe der vier beteiligten Erregungen berechnet werden:
![]() .
.
Setzt man für die vier Radiusvektoren r1 bis r4 die hergeleiteten Beziehungen ein und fasst die gemeinsamen Faktoren zusammen, so ergibt sich folgendes:
![]()
Für die Feuerraten f1 und wählten wir bereits den exponentiellen Ansatz gemäß
![]()
![]()
Hierbei beschreibt die Urgröße u beispielsweise dem Gelenkwinkel beim Heben des Armes nach vorn.
Analog wählten wir für die zwei übrigen Feuerraten f2 und f4, die das seitliche Bewegen des Armes regeln, die Vorgabe
![]()
![]()
Setzen wir die obigen Feuerraten ein, so ergibt sich für die Erregung eines Outputneurons im Punkt P(x,y) die Funktionsgleichung
![]()
Die
Verwendung der Hyperbelfunktion ![]()
liefert die Vereinfachung
![]()
Nun wenden wir folgendes Additionstheorem für Hyperbelfunktionen an
![]()
und erhalten
![]()
Wegen
![]()
![]()
Gilt nun
![]()
Diese Erregungsfunktion besitzt ein globales Maximum, da ihre Hessematrix im Überlagerungsgebiet der Ursprungsfunktionen f1 bis f4 negativ definit ist.
An der Stelle des Maximums verschwinden die partiellen Ableitungen nach x und y. Daher berechnen wir diese Ableitungen und setzen sie gleich Null.
![]()
Wir untersuchen nur den Faktor F2
![]()
Nur der Faktor F2 kann gleich Null werden, denn die mittlere Feuerrate fm ist größer als Null, die Längskonstante λ ebenfalls, die Funktion cosh(x) ist immer positiv und auch die Exponentialfunktion ist immer größer als Null.
Daher ergibt sich aus dem Nullsetzen der partiellen Ableitung nach x die Extremwertbedingung
![]()
Auflösen nach x liefert die Gleichung
![]()
![]()
![]()
Für u + v leiten wir eine ähnliche Formel her, indem wir die partielle Ableitung nach y gleich Null setzen.
![]()
mit
Die Ableitung nach y kann nur Null werden, wenn F3 gleich null wird.
![]()
Daraus ergibt sich analog die Nullstellenbedingung
![]() −v
einmal addieren und einmal subtrahieren, kommen wir auf die
Bestimmungsgleichungen für u und v.
−v
einmal addieren und einmal subtrahieren, kommen wir auf die
Bestimmungsgleichungen für u und v.
![]()
![]()
Damit haben wir die Bedingungen für die Parameter u und v erhalten, die gewährleisten, dass die Erregungsfunktion f(x;y) im Punkt P(x;y) des Cortex ein globales Maximum annimmt.
Indirekt haben wir auch eine Forderung an die Variablen x und y erhalten. Die Funktion artanh ist nur definiert für Werte im Intervall von -1 bis +1. Diese Bedingung müssen x und y erfüllen.
Nun sollten wir uns einen Überblick über die hergeleiteten Ergebnisse verschaffen. Wir erinnern uns an das Diagramm der Urgrößen u und v. Hier war der Radiusvektor t und der Winkel ω zugeordnet. Diesen Winkel können wir berechnen. Wir erinnern uns:
![]()
![]()
Einsetzen von v und u liefert die Gleichung
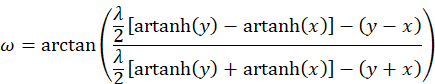
![]()
Eine einfachere Formel für den Phasenwinkel ω ergibt sich, wenn wir u und v durch den Radius t und den Phasenwinkel ausdrücken. Dann gilt für die Summe und die Differenz von u und v die Beziehung
![]()
![]()
Dann gilt auch
![]()
Wegen
![]()
![]()
Gilt deshalb
![]()
Umgestellt ergibt sich für den Phasenwinkel ω die einfachere Gleichung
![]()
Für kleine Werte ist die Funktion artanh(x) näherungsweise gleich x.
![]()
Daher gilt für kleine x und y auch die Näherung
![]()
Drückt man x und y durch den Radiusvektor r und den Winkel α aus, so ergibt sich folgendes:
![]()
![]() für |x| << 1
für |x| << 1
Zwischen dem Phasenwinkel ω zum Radiusvektor t der zwei Urgrößen u und v und dem Phasenwinkel α vom Radiusvektors r im cortikalen Divergenzgebiet besteht also eine Phasenverschiebung von π/4 für kleine Werte von x und y, hier läuft der Winkel ω dem Winkel α um etwa 45 Grad hinterher.
Der Zeiger r, der zum aktuellen Erregungsmaximum der cortikalen Erregungsfunktion zeigt, ist um π/4, also 45 Grad verschoben gegen den Phasenwinkel ω der Urgrößen, wenn die Werte x und y klein sind. Rotiert der Vektor t, so rotiert auch der Vektor r, beide im gleichen Rotationssinn, jedoch leicht um circa 45 Grad verschoben. Der Urgrößenzeiger hinkt dem Zeiger r um etwa 45 Grad hinterher.
Nun wird klar, warum eine maximalerregte Neuronenpopulation um ein Zentrum rotiert, wenn der Affe mit seiner Hand gleichfalls eine Rotation um ein Zentrum ausführt.
Die Identität
![]()
ergibt durch Einsetzen der Gleichungen für die Summe und die Differenz von u und v die Gleichung
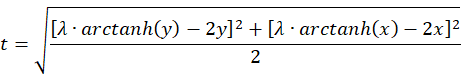
Wir prüfen nach, wie eine Näherung für kleine Werte von x und y aussieht, bei der die Funktion artanh(x) ungefähr gleich x ist.
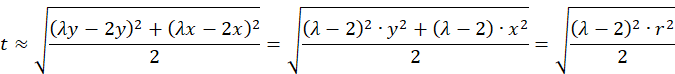
![]() für
kleine r, also für kleine x und kleine y.
für
kleine r, also für kleine x und kleine y.
Speziell beim Wert
gilt für kleine r die Näherungsformel
![]()
Die zwei Radiusvektoren t und r
sind für den Wert ![]() und
kleine Werte von r und t näherungsweise gleich groß, außerdem bilden sie dann
den Winkel von π/4 (also 45 Grad) zueinander, der Radiusvektor t läuft dem
Radiusvektor r um 45 Grad verschoben hinterher.
und
kleine Werte von r und t näherungsweise gleich groß, außerdem bilden sie dann
den Winkel von π/4 (also 45 Grad) zueinander, der Radiusvektor t läuft dem
Radiusvektor r um 45 Grad verschoben hinterher.
Am Ende wollen wir den Radiusvektor t und den Phasenwinkel ω in Abhängigkeit von den Größen x und y graphisch in Diagrammen darstellen. Zur Erinnerung: x und y sind die Koordinaten des Extremwertes, des Erregungsmaximums in der x-y-Ebene, während t und ω die Urgrößen u und v in Polarkoordinaten beschreiben.
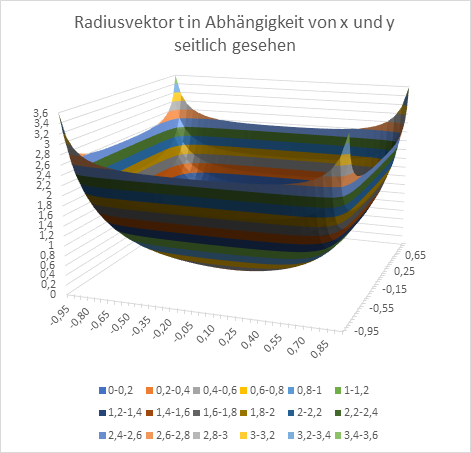
Abbildung 16: Radiusvektor t in Abhängigkeit von x und y - seitlich gesehen
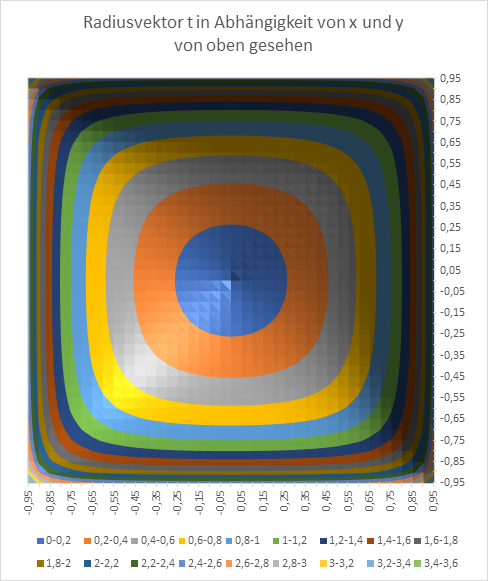
Abbildung 17: Radiusvektor t in Abhängigkeit von x und y - von oben gesehen
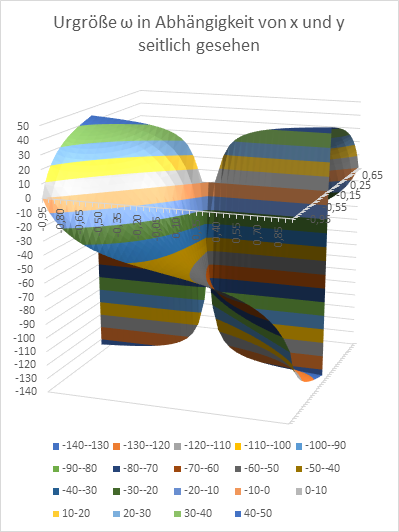
Abbildung 18: Urgröße w in Abhängigkeit von x und y - seitlich gesehen
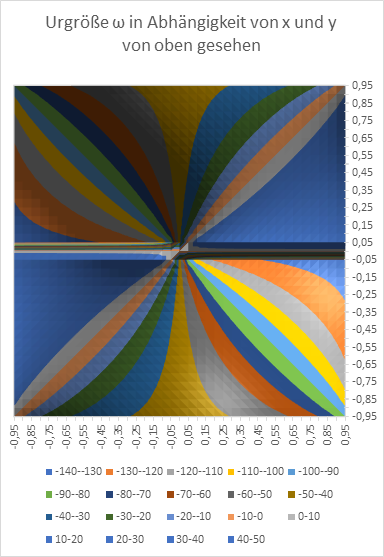
Abbildung 19: Urgröße w in Abhängigkeit von x und y - von oben gesehen
Das obige Diagramm erinnert stark an die Darstellungen zu den Orientierungssäulen im primären visuellen Cortex. Dies liegt daran, dass auch dort ein Divergenzmodul verwendet wird. Anstelle von Muskelspannungen werden jedoch visuell bewertbare Urgrößen verwendet. Das Prinzip aber bleibt das Gleiche.
Mathematiker werden sich etwas wundern. Sie berechnen die Lage eines Extremwertes in Abhängigkeit von den Parametern einer Funktion. Wir gehen den umgekehrten Weg. Aus der Lage des Extremwertes berechnen wir die ursprünglich dazu gehörenden Parameter, z. B. die Gelenkwinkel oder die zugehörigen Feuerraten der Muskelspannungssensoren. Denn wir wollen wissen, was die cortikalen Erregungsmaxima in den primären Cortexfeldern für eine Bedeutung haben.
Die primären Cortexgebiete (sie entsprechen den sensorischen Zentren des alten Strickleiternervensystems) projizieren zum motorischen Cortex (die den motorischen Zentren entsprechen) - ohne die Feuerrate zu verändern. Auch die Projektionsaxone, die dies bewirken, vermischen sich nicht, sondern übertragen die Topologie in die Zielgebiete.
Deshalb messen Neurologen im motorischen Cortex die Bewegung von maximal erregten Neuronenpopulationen bei motorischen Aktivitäten. Der Output der sensorischen Seite findet sein Ebenbild in der motorischen Seite.
Und wir erhalten ein erstaunliches Resultat. Die Orte der maximalen Erregung in den primären motorischen Cortexgebieten verschlüsseln die Gelenkwinkel. Wir können erklären, warum die Erregungsmaxima im Cortex hin- und herwandern und was sie im motorischen Fall bedeuten.
Die Praxistests bestätigen diese Theorie. Bewegt ein Affe seinen Arm kreisförmig hin- und her, so wandert das Erregungsmaximum im Cortex ebenfalls kreisförmig hin- und her. Die experimentellen Messungen bestätigen diese Theorie der neuronalen Divergenzmodule.
Da etwa ein Drittel der Cortexrinde zu den primären Gebieten gezählt wird, können wir mit dieser Theorie die Funktionsweise eines Drittels der Cortexrinde erklären. Und dies, ohne auch nur eine einzige Synapse untersucht zu haben. Dies mögen insbesondere die Wissenschaftler des Human Brain Projekts zur Kenntnis nehmen. Sie analysierten Millionen von Synapsen, ohne die Funktionsweise der Cortexrinde erklären zu können.
Die Leistungsfähigkeit der Divergenzmodule im Gehirn wird nachfolgend auf die primären visuellen Cortexgebiete angewendet.
4.2.2. Das Helligkeitsmodul mit seitlicher Signalüberlagerung
Im primären visuellen Cortex beobachtet man sogenannte Orientierungssäulen. Deren Neuronen reagieren besonders empfindlich auf visuelle Stimuli. Wird im Blickfeld eine geneigte, dunkle Gerade vor einem hellen Hintergrund gezeigt, so reagieren bestimmte Populationen von Neuronen mit einem starken Anstieg der Feuerrate. Diese Populationen von Neuronen sind windmühlenartig um ein gemeinsames Zentrum angeordnet. Jedem Anstiegswinkel entspricht eine andere Population mit maximaler Erregung.
Immer dann, wenn Erregungsmaxima auftreten, oder wenn sie anfangen, sich zu bewegen und ihre Lage verändern, sollten bei Mathematikern die Alarmglocken läuten. Auch ich vermutete vor vielen Jahren, dass es sich hier um ein Extremwertproblem handelt. Sollten meine Kenntnisse aus der Integral- und Differentialrechnung es ermöglichen, dieses Phänomen der visuellen Orientierungssäulen auf naturwissenschaftliche Füße zu stellen? Schon Professor Günther Porath lehrte mich während meines Studiums in Güstrow vor über 50 Jahren, dass die Mathematik kein Selbstzweck sei, und dass es jeden Mathematiker mit Freude und Stolz erfüllen sollte, wenn er ein neues mathematisches Anwendungsgebiet erschließen konnte. Sollte uns die Differential- und Integralrechnung einen Zugang zum Verständnis der Funktionsweise der Orientierungssäulen in V1 eröffnen?
Können uns die Fähigkeiten, Extremwertprobleme zu lösen, auch bei den primären visuellen Feldern zu neuen Einsichten führen?
Die Antwort lautet eindeutig ja. Die Winkelempfindlichkeit der Orientierungssäulen im primären visuellen Cortex ist ein Ergebnis des Wirkens von elementaren Naturgesetzen. Dies wird nachfolgend bewiesen und wurde bereits in meinen früheren Veröffentlichungen dargestellt.
Wichtig ist insbesondere, dass zur Entstehung der Winkelempfindlichkeit der Orientierungssäulen keinerlei Lernvorgänge nötig sind. Man braucht kein neuronales Netz, welches mit Millionen von Datensätzen gefüttert werden muss, und welches den Output ständig mit irgendwelchen Vorgaben (wo kommen die her?) vergleicht und anpasst. Diese Winkelempfindlichkeit wird nicht erlernt, sie ist einfach da, sobald die neuronale Schaltung des zugehörigen Divergenzmoduls vollständig ausgereift ist. Bei Fliegen etwa direkt nach dem Schlüpfen. Die Lebensdauer einer Fliege wäre auch nicht ausreichend, um ein neuronales Netz durch Erfahrung zu programmieren. Manche Fliegenarten leben nur einen Tag lang.
Und dies ist der springende Punkt! Die Hirnforscher des Human Brain Projekts sind offenbar daran interessiert, lernende neuronale Netze im Cortex zu finden. Dazu studieren sie die fast unendlich große Zahl der synaptischen Verbindungen und erinnern sich daran, dass der weltbekannte Nobelpreisträger Eric Kandel nachweisen konnte, wie die Veränderung der synaptischen Stärke zu Lernvorgängen führt. So hoffte man, die Entstehung von Intelligenz und Denken als Resultat des Lernens in neuronalen Netzen zu entlarven.
Hierbei ist die Hoffnung die Triebfeder. So wurden alle verfügbaren Ressourcen der Europäischen Union in dieses Ziel gesteckt und hochfliegende Pläne entworfen. Alle Skeptiker - oft ebenfalls hochdekorierte Hirnforscher - wurden mehr oder weniger genötigt, sich in dieses Projekt zu integrieren, da es nur noch dann dringend benötigte Fördermittel gab, wenn man sich an der Sache beteiligte.
Damit war jedoch die Freiheit der Forschung in Gefahr!
Was wollen die Verantwortlichen dieses Projekts entgegnen, wenn man sie auffordert, folgende Fakten zur diskutieren:
1.Die Cortexrinde ist fast überall myelinfrei, insbesondere die Schicht 4.
2.Die Schicht 4 empfängt in den primären Gebieten den Input der Rezeptoren.
3.Sie übergibt den Input an Outputneuronen, deren Anzahl die Anzahl der Inputneuronen um ein Vielfaches übertrifft.
4.Die Ausbreitung der Inputerregung zu den Outputneuronen führt zu lokalen Erregungsmaxima, erst diese ermöglichen es den Hirnforschern, einer bestimmten Gehirnaktivität eine Ursache zuzuschreiben (Bsp.: Broca-Zentrum-Sprache).
5.Erregungsmaxima erfordern eine konkave Übertragungsfunktion.
6.Die mathematische Modellierung des Inputs durch Parameter (Gelenkwinkel, Tonhöhe, visuelle Stimuli, -) macht es möglich, die Theorie der Extremwertbestimmung anzuwenden und so Zusammenhänge zwischen Parametern und dem Ort der Maximalerregung herzuleiten.
7.Das Auftreten dieser Erregungsmaxima in den primären Cortexgebieten ist keine Folge des Wirkens von neuronalen Netzen, sondern eine Folge der Signaldämpfung bei der Ausbreitung der Erregung in der Ebene.
8.Mindestens ein Drittel der Cortexgebiete sind primäre Gebiete, so dass sich auf diese Weise etwa ein Drittel der Gehirnfunktionen erklären lassen.
Besteht man dann immer noch auf einem Alleinvertretungsanspruch auf dem Gebiet der Hirnforschung?
Die Hirnforscher dieser Welt sollten sich vom Human Brain Projekt nicht mehr auf der Nase herumtanzen lassen!
Nachfolgend wird die Winkelempfindlichkeit der visuellen Orientierungssäulen mathematisch hergeleitet. Es wird eine andere Variante der Herleitung gewählt als in meiner vorangegangenen Monografie, die Ergebnisse sind jedoch die Gleichen.
Teile des Textes wurden teils unverändert aus meiner Monografie -Gehirntheorie der Wirbeltiere- übernommen, ohne sie als Zitate zu kennzeichnen. Es handelt sich bei dieser Arbeit auch nicht um eine Dissertation, so dass Plagiatsjäger ihre Energie nicht darauf verschwenden sollten.
Im visuellen Cortex V1 der Wirbeltiere beobachtet man eine Richtungsselektivität der sogenannten Orientierungssäulen. Dies sind Neuronengruppen, die sich durch alle sechs Schichten des visuellen Cortex ziehen und kleine zylinderförmige Säulen bilden. Deren Aktivität ist selektiv empfindlich auf den Neigungswinkel einer dunklen Linie vor hellem Hintergrund. Die beteiligten Neuronen müssen also auf dunkle Objekte ansprechen, sie sind daher vom Dunkel-On-Typ.
Der Input des zweiten Segments kommt aus der Retina. Und bereits dort hat sich ab einem gewissen Stand der Evolution eine Neuronengruppe mit großen dendritischen Feldern etabliert. Ihre Signale treffen im sensorischen Kern des zweiten Segments ein, der auch als visueller Thalamus bezeichnet wird, und bilden dort eine Schicht mit magnocellularen Input.
Über aufwärts projizierende Neuronen der Klasse 4 erreichen diese Signale das sensorische Zentrum des ersten Segments, welches den visuellen Cortex darstellt. Denn bereits im Thalamus erfolgte die Aufspaltung der Modalitäten, jede zog in eine eigene Modalitätenleiter des Strickleitersystems. Aus diesen Modalitätenleitern der ersten Etage entstanden die verschiedenen Lobi des Gehirns. Im Occipilallobus endeten die visuellen Signale in der vierten Cortexschicht. Somit bestand die vierte Schicht des visuellen Cortex aus einer unteren Teilschicht, die als Input die magnocellularen Signale aus dem visuellen Thalamus empfing. Diese dienten dem Hell-Dunkel-Sehen.
Wir analysieren zunächst die Reaktion einer magnocellularen Ganglienzelle der Retina vom Typ Dunkel-On, wenn über ihr rezeptives Feld eine dunkle Linie vor hellem Hintergrund mit einem konstanten Anstellwinkel von links nach rechts bewegt wird. Die Linie wird quasi parallel zu sich selbst verschoben. Wir denken uns das rezeptive Feld der Ganglienzelle durch einen Kreis dargestellt. Jedes dunkle Objekt im Kreis wird eine Erregung der Ganglienzelle bewirken, so auch diese dunkle Gerade.
Für die Erkennung heller Linien auf dunklem Untergrund sind (nach Ansicht des Autors) die Hell-On-Ganglienzellen zuständig. Wir konzentrieren uns hier auf die Dunkel-On-Zellen.
Die Gerade möge den Kreis schneiden, der das rezeptive Feld der Zelle darstellt, so dass ein Teil von ihr zur Sehne im Kreis wird. Die Länge dieser Sehne wird die Feuerrate der Ganglienzelle bestimmen. Wir gehen hier davon aus, dass die Erregung der Zelle mit dem Quadrat der Sehnenlänge zunimmt. Später können wir diese Voraussetzung möglicherweise wieder fallenlassen, wenn wir das Wirkprinzip der Orientierungssäulen begriffen haben. Zunächst interessieren wir uns nur für die Sehnenlänge.
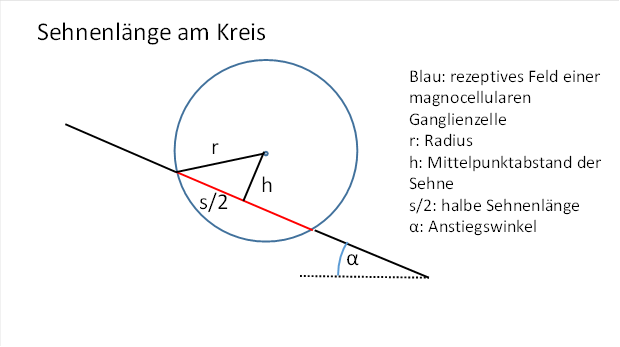
Abbildung 20: Sehnenlänge am Kreis
Die Sehnenlänge wird von der Lage der Geraden abhängen, genauer vom Abstand der Geraden zum Kreismittelpunkt.

Abbildung 21: Veränderung der Sehnenlänge bei paralleler Verschiebung zu sich selbst
Eine geeignete Größe zur Berechnung der Sehnenlänge ist der Abstand der Sehne vom Kreismittelpunkt.
Der bekannte Radius r bildet die Hypotenuse eines rechtwinkligen Dreiecks, dessen Katheten h und die halbe Sehnenlänge s/2 sind. Diese drei Größen sind über den Satz des Pythagoras miteinander verknüpft.
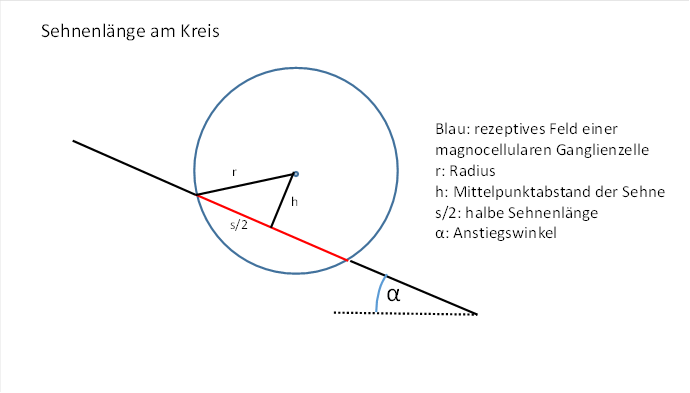
Abbildung 22: Berechnung der Sehnenlänge am Kreis
Es gilt also: ![]() (Pythagoras)
(Pythagoras)
Daraus ergibt sich für das Quadrat der Sehnenlänge folgende Gleichung:
![]()
Generell muss
![]() gelten, da r die Länge der
Hypotenuse eines rechtwinkligen Dreiecks entspricht und h die Länge einer
Kathete darstellt. Anderenfalls wird das Quadrat der Sehnenlänge negativ, also
die Sehnenlänge eine imaginäre Zahl. Dann schneidet die Gerade den Kreis nicht
mehr, die Formel wird nicht mehr anwendbar.
gelten, da r die Länge der
Hypotenuse eines rechtwinkligen Dreiecks entspricht und h die Länge einer
Kathete darstellt. Anderenfalls wird das Quadrat der Sehnenlänge negativ, also
die Sehnenlänge eine imaginäre Zahl. Dann schneidet die Gerade den Kreis nicht
mehr, die Formel wird nicht mehr anwendbar.
Wir gehen hier davon aus, dass die Feuerrate f der Ganglienzelle, in deren rezeptiven Feld die Sehne mit der Länge s das Gesichtsfeld verdunkelt, quadratisch mit der Sehnenlänge zunimmt. Einen möglichen Proportionalitätsfaktor werden wir hier vernachlässigen, da er bei Extremwertaufgaben keine Wirkung hat.
![]()
Wir betrachten nun die Sehnenlänge bzw. ihr Quadrat unter Verwendung eines Koordinatensystems, bei dem sich das rezeptive Feld nicht im Koordinatenursprung befindet. Der Mittelpunkt des Kreises K, der das rezeptive Feld der Ganglienzelle darstellen soll, möge im Koordinatensystem der Punkt Pk sein, dem wir die Koordinaten xm und ym zuordnen. Dann können wir den Punkt Pk auch als Pk(xm; ym) darstellen.
Die Gerade g, die den Kreis K schneiden möge, habe den Anstellwinkel φ zur x-Achse. Ihr Abstand vom Koordinatenursprung P(0;0) sei d.
Der Kreismittelpunkt Pk(xm; ym) möge vom Koordinatenursprung P(0; 0) den Abstand rk besitzen. Die Gerade, die beide Punkte verbindet, hat also die Länge rk. Sie möge mit der x-Achse den Winkel φk bilden.
Zur
Herleitung wird vorwiegend der Satz des Pythagoras verwendet sowie die
Darstellungen der Größen xm, ym über den Sinus und
Cosinus der Winkeldifferenz![]() .
Nützlich ist die grafische Darstellung der zur Herleitung nötigen Zusammenhänge
in nachfolgender Abbildung.
.
Nützlich ist die grafische Darstellung der zur Herleitung nötigen Zusammenhänge
in nachfolgender Abbildung.
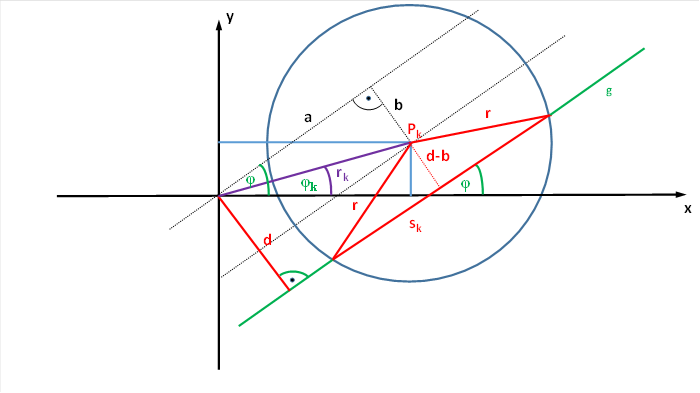
Abbildung 23: Herleitung der Fromeln für die Sehnenlänge rezeptiver Felder
Die Gerade g schneide die x-Achse im Winkel φ. Der Kreismittelpunkt habe vom Koordinatenursprung den Abstand rk. Die Strecke rk bilde mit der x-Achse den Winkel φk.
Die Gerade g, die das rezeptive Feld des Neurons, also den Kreis schneidet, habe vom Koordinatenursprung den Abstand d. Ihr Abstand vom Kreismittelpunkt sei gleich d-b.
Die Gerade schneide den Kreis an zwei Stellen, wodurch die Sehne sk entsteht.
Dann können wir die Länge der halben Sehne über den Satz des Pythagoras berechnen.
![]()
Die Größe b ergibt sich über den Sinus der Winkeldifferenz der zwei beteiligten Winkel.
![]()
Somit gilt
![]()
Eingesetzt in die Gleichung für die halbe Sehnenlänge ergibt sich letztlich die Gleichung
Wir gehen (vereinfachend) davon aus, dass die Feuerrate gleich dem Quadrat der Sehnenlänge sk ist.
Nachdem wir eine Formel für die Feuerrate einer magnocellularen Ganglienzelle mit einem großen rezeptiven Feld hergeleitet haben, betrachten wir vier benachbarte Ganglienzellen der Retina. Wir ordnen ihnen Feuerraten f12, f2, f3 und f4 zu und stellen uns vor, ihre Signale mögen über den Sehnerv und den visuellen Thalamus den primären visuellen Cortex erreichen. Dabei mögen ihre Feuerraten unverändert bleiben, auch die räumliche Anordnung zueinander möge erhalten bleiben, weil die Abbildung der Netzhaut im Cortex topologietreu ist.
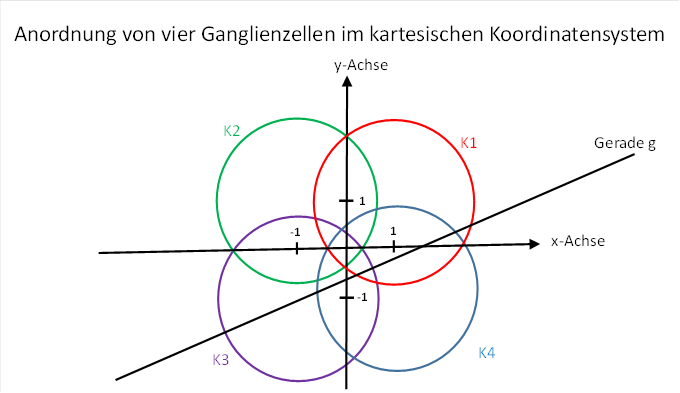
Abbildung 24: Anordnung von vier Ganglienzellen im kartesischen Koordinatensystem
Jede der vier Ganglienzellen habe bezüglich der Retina ein kreisförmiges rezeptives Feld mit dem Radius rs. Wichtig ist die teilweise oder völlige Überlappung der vier rezeptiven Felder. Die Gerade habe wieder den Anstiegswinkel φ.
Die
Mittelpunkte der vier Kreise sollten von der x-Achse und von der y-Achse
jeweils den Abstand 1 besitzen. Damit haben sie vom Koordinatenursprung den
Abstand ![]() .
.
Bei dieser Anordnung der Ganglienzellen gelten folgende Besonderheiten:
![]()
![]()
![]()
![]()
![]()
Damit vereinfachen sich die Formeln für die Sehnenlängen s1 bis s4 und damit für die Feuerraten f1 bis f4.
![]()
![]()
![]()
![]()
Wir vereinfachen.
Es gilt
![]()
![]()
![]()
Einsetzen in die Feuerraten liefert die endgültigen Gleichungen.
![]()
![]()
![]()
Die vier Feuerraten f1 bis f4 werden über den Sehnerv von den Ganglienzellen zum visuellen primären Cortex übertragen und erregen vier Inputneuronen in der vierten Cortexschicht. Die zugehörigen Outputneuronen befanden sich in der direkt benachbarten Schicht 3.
Doch auch damals waren Neuronen störanfällig, sie konnten ausfallen. Günstig waren dann Ersatzneuronen in der Nähe, die die Signale ebenfalls empfangen konnten. Die Anzahl der Outputneuronen musste daher anwachsen. Die Outputschicht wuchs in die Breite. Die Abstände der Inputneuronen nahmen dabei zu, denn sie mussten nun viel mehr Outputneuronen erregen. Die Reserveneuronen, die einen Ausfall kompensieren konnten, wurden bereits -fest verdrahtet-, besaßen also alle nötigen Synapsen. So musste bei einem Neuronenausfall nicht erst ein Ersatzneuron gebildet werden.
Genau diese Strategie führte zur Herausbildung von Neuronenpopulationen, die wir gemeinhin als Orientierungssäulen bezeichnen.
Wir betrachten die Schicht S4. Sie empfange den Input der großen Dunkel-On-Neuronen der Retina. Wir konzentrieren uns auf die vier magnocellularen Ganglienzellen der Retina, deren vier rezeptive Felder je nach der Neigung einer dunklen Geraden vor hellem Hintergrund die bereits berechneten Feuerraten f1 bis f4 generieren. Diese vier Feuerraten treffen - ohne Abschwächung - über den Thalamus auf den myelinisierten im primären visuellen Cortex in der Schicht 4 ein. Sie erregen dort vier Inputneuronen N1 bis N4.
Wir stellen diese ebenfalls symbolisch in einem Koordinatensystem dar. Die bisherige Anordnung der Ganglienzellen möge auf die vier Inputneuronen N1 bis N4 übertragen werden.
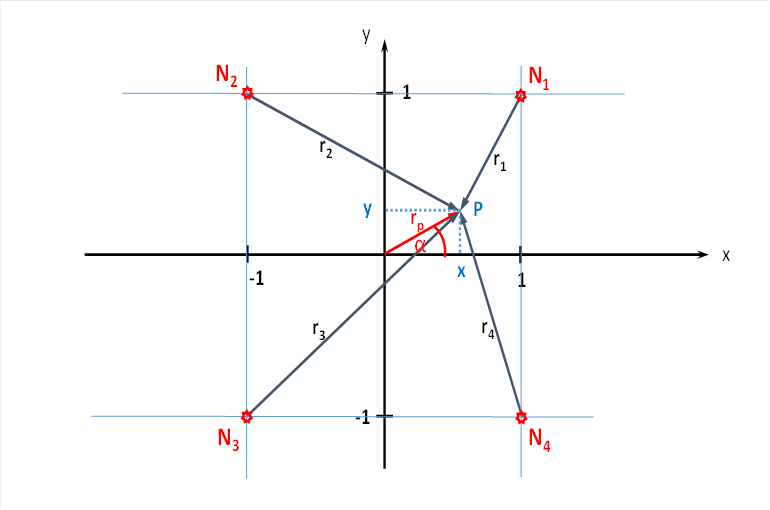
Abbildung 25: Vier cortikale Outputneuronen im Helligkeitsmodul mit seitlicher Signalüberlagerung
Die im visuellen Cortex eintreffenden Signale werden von den vier Inputneuronen zu den unzähligen Outputneuronen der Schicht 5 übertragen, wobei die Hilfe der Interneuronen der Schicht 4 zu Hilfe genommen wird. Die Ausbreitung erfolgt allseitig in der Fläche und genügt der Kabelgleichung für marklose Fasern. Diese Kabelgleichung müssen wir wegen der Erregungsausbreitung in der Fläche etwas modifizieren. Im Exponenten tritt das Quadrat des Abstandes auf.
Wir untersuchen nun, welche Erregung ein Outputneuron im Punkt P mit den Koordinaten x und y erhält. Die Strecke zwischen dem Koordinatenursprung und dem Punkt P bezeichnen wir als Radiusvektor rp, dieser bildet mit der x-Achse den Winkel α. Dann können wir später x und y durch rp und den Sinus beziehungsweise den Kosinus des Winkels α ausdrücken.
Das Outputneuron im Punkt P empfängt von jedem der vier Inputneuronen einen Erregungsanteil, die Gesamterregung ergibt sich durch einfache Addition.
![]()
Wir verwenden den Index E bei der Feuerrate f, weil wir darauf hinweisen wollen, dass die Erregungsausbreitung in der Ebene erfolgt.
Wir werden diese Funktion als Erregungsfunktion fE(x,y) des Helligkeitsmoduls mit seitlicher Signalausbreitung bezeichnen.
![]()
Sie wird uns im Helligkeitsmodul mit räumlicher Signalausbreitung wiederbegegnen.
Wir gehen davon aus, dass das Outputneuron im Punkt P Bestandteil einer Orientierungssäule des visuellen Cortex ist. Bei einem bestimmten Anstellwinkel φ der Geraden g erreicht die Erregung dieses Outputneurons ein Maximum. Wir wollen ergründen, unter welchen Bedingungen dies geschieht.
Wir müssen also ermitteln, wann die Feuerrate f ein Maximum annimmt. Dies ist der Fall, wenn die Ableitung der Funktion f gleich Null wird. Aber nach welcher Größe müssen wir die Ableitung bilden?
Das Outputneuron ist ortsunveränderlich. Es wandert nicht im Cortex hin und her. Daher sind die Größen x und y konstant.
Im Versuch zur Ermittlung der Richtungsselektivität der Orientierungssäulen wurde eine Gerade parallel zu sich selbst verschoben. Damit ist der Anstellwinkel φ ebenfalls eine Konstante. Die einzige veränderliche Größe in der Versuchsanordnung ist der Abstand d der Geraden vom Koordinatenursprung. Dieser ändert sich, wenn die Gerade zu sich selbst parallelverschoben wird.
Wir müssen also die Funktion f nach der Größe d differenzieren und beachten, dass alle anderen Größen Konstante sind.
Die Ableitung einer Summe ist gleich der Summe der Ableitung der einzelnen Summanden. Die exponentiellen Größen sind ebenfalls Konstante.
![]()
Für die Feuerraten hatten wir bereits Formeln hergeleitet:
![]()
![]()
![]()
![]()
Wir berechnen daher zunächst die Ableitungen der Feuerraten f1 bis f4 nach d, um sie später zu addieren.
![]()
![]()
![]()
Diese Werte setzen wir ein und setzen die Ableitung gleich Null, um die Bestimmungsgleichung für das Vorliegen eines Maximums zu ermitteln. Da alle Komponenten konkave Funktionen sind, existiert im gemeinsamen Konkavitätsgebiet ein globales Maximum. Durch Nullsetzen der Summe der Ableitungen erhalten wir eine Bedingungsgleichung für das Maximum von f.
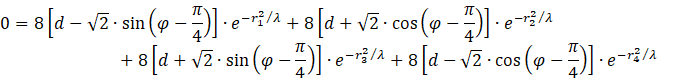
Der Faktor 8 kann herausdividiert werden. Ausmultiplizieren und Zusammenfassen ergibt die Gleichung
![]()
![]()
Für die Größen r1 bis r4 haben wir im letzten Kapitel bereits folgende Gleichungen hergeleitet
![]()
![]()
![]()
![]()
Wir setzen zur Verkürzung der Formeln
![]()
![]()
![]()
So erhalten wir die Extremwertgleichung
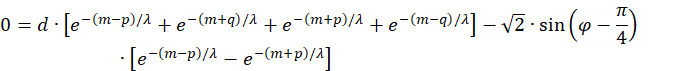
![]()
Der allen Summanden gemeinsame
Faktor ![]() kann
herausdividiert werden, er ist immer größer als Null.
kann
herausdividiert werden, er ist immer größer als Null.
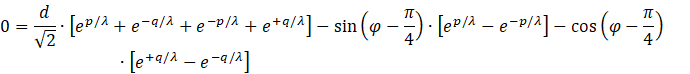
Die Anwendung der Definitionsgleichungen
![]()
![]()
ergibt
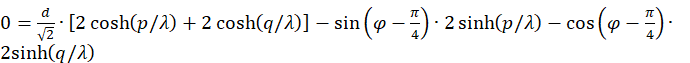 .
.
Auf den Faktor 2 können wir verzichten.
Diese Gleichung muss erfüllt sein, damit das Neuron im Punkt P maximal feuert, also dort ein Erregungsmaximum auftritt. Wir wenden ein Additionstheorem an.
![]()
Damit gilt auch
![]() .
.
Wegen
![]()
![]()
Gilt dann
![]()
![]()
Wir wenden folgende Additionstheoreme für Summen von Sinus und Cosinus an
![]()
![]()
![]()
![]()
Nun können wir ein altbekanntes Additionstheorem für Winkelfunktionen anwenden.
![]()
Dann erhalten wir eine neue Gleichung
![]()
![]()
Wir lösen nach dem Winkel φ auf.
![]()
![]()
Diesen Winkel φ muss die Gerade besitzen, damit die Feuerrate des Outputneurons im Punkt P(x,y) ein Maximum annimmt, wenn der Abstand der Geraden vom Koordinatenursprung den Wert d besitzt.
Damit haben wir endgültig den Winkel φ ermittelt, bei dem das cortikale Outputneuron im Punkt P ein globales Erregungsmaximum annimmt.
Wir sehen, dass dieser Winkel eine periodische Funktion der Neuronenkoordinaten x und y darstellt und außerdem vom Nullpunktabstand der geneigten Geraden mit dem Anstellwinkel φ abhängt.
Wir können diese Abhängigkeit für kleine Werte von x und y näherungsweise ermitteln.
Wegen tanh(x) ≈ x für kleine x gilt die Näherung
![]()
![]()
Die Größen rp und α finden wir in der Koordinatendarstellung des visuellen Cortex wieder. Weitere Umformung ergibt für die Näherungsformel (für kleine y und y) die Darstellung
![]()
![]()
Wegen
![]()
Gilt auch
![]()
Daher gilt die für die Näherungsformel für kleine x, y und somit auch rp die Vereinfachung
Da jedoch
![]()
ist, nimmt für kleine rp die Näherungsformel folgende Gestalt an:
![]()
Für kleine x, y und rp gibt es zwischen dem Anstellwinkel φ der geneigten Geraden im Sehfeld und dem Radiusvektor rp im visuellen Cortexfeld eine Phasenverschiebung von 1-π/2, also -32 Grad.
Zuletzt wollen wir den Zusammenhang zwischen x, y und dem Anstellwinkel φ für den Fall grafisch analysieren, dass das betreffende Outputneuron maximal erregt wird. Die ist für wechselnde x und y jeweils verschieden. In einem zweidimensionalen Excel-Diagramm lässt sich das gut erkennen.
Die Originalformel lautete:
![]()
Wichtig ist es, darauf hinzuweisen, dass die Funktion arcsin(x) nur für Werte mit |x|<1 definiert ist. Dies ist nur dann erfüllt, wenn die geneigte Gerade die Kreise K1 bis K4 tatsächlich schneidet. Nur dann ist die Sehnenlänge eine reelle Zahl. Schneidet die Gerade die Kreise nicht, so gibt es auch keine Feuerraten, die vier beteiligten magnocellularen Ganglienzellen in der Retina liefern dann kein Signal zum Cortex. Die Formel für den Anstellwinkel φ der geneigten Geraden ist dann nicht anwendbar.
Wir zeigen nachfolgend das x-y-Diagramm, welches mit Excel erzeugt wurde. Hier wurde als Nullpunktabstand der geneigten Geraden der Wert d = 0,1 verwendet. Damit die Erregung im Punkt x und y den Maximalwert annimmt, mit der Anstellwinkel φ der Geraden die im Diagramm ersichtlichen Werte annehmen (im Bogenmaß).
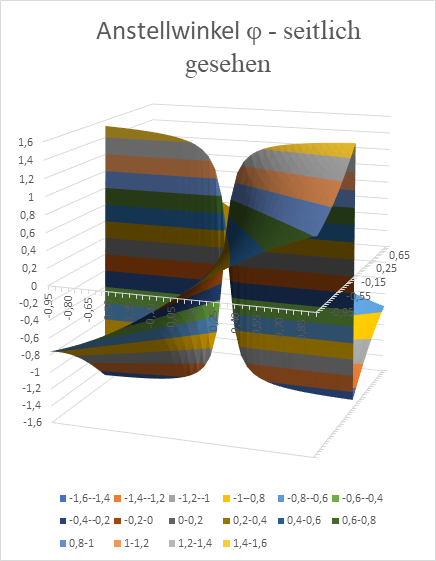
Abbildung 26: Anstellwinkel Phi einer Geraden als Funktion von x und y - seitlich gesehen
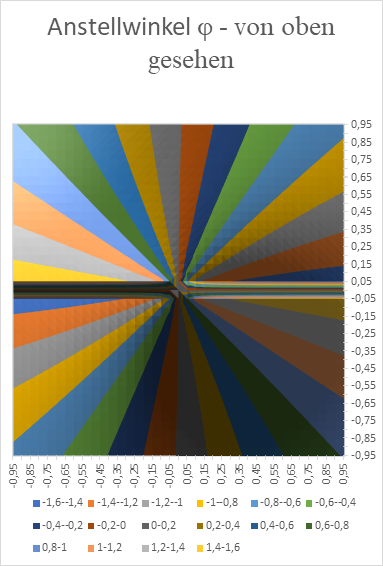
Abbildung 27: Anstellwinkel Phi im Helligkeitsmodul mit seitlicher Signalüberlagerung - von oben gesehen
Wir sehen die typische windmühlenartige Anordnung der Neuronenpopulationen für die verschiedenen Anstellwinkel φ der Geraden.
Die laterale Nachbarhemmung führt übrigens im Zusammenwirken mit dem -Wettlauf der Aktionspotentiale- dazu, dass nur Outputneuronen in unmittelbarer Nähe vom Erregungsmaximum erregt sind. Diese Neuronen erreichen ihr Aktionspotential am schnellsten, feuern also als erste. Ihr Aktionspotential vernichtet die Erregung aller Neuronen des Umfeldes, da diese Neuronen die Erregungsschwelle für die Auslösung eines Aktionspotentials noch nicht erreichen konnten. Der Neurologe sagt dazu: The Winner Takes all it. Der Gewinner bekommt alles!
Dieser frühe Urzustand des magnocellularen visuellen Systems wurde verändert durch ein Dickenwachstum der Schicht 3. Zuvor entstanden die magnocellularen Ganglienzellen vom Hell-On-Typ. Der Hell-On-Typ war mit dem Dunkel-On-Typ signalkompatibel.
Dadurch konnte die Outputschicht des Dunkel-On-Typs auch vom Input aus der darüberliegenden Schicht 4-Hell-On mitversorgt werden. Als ein Effekt trat nun die Bildung von Orientierungssäulen auf, die nicht nur auf schwarze oder weiße Linien reagierten, sondern auf Linien mit beliebigen Grauwerten vor einem grauen Hintergrund mit anderem Grauwert.
Genau wie beim Farbmodul mit vertikaler Signalausbreitung fand nun, als die Outputschichten der Cortexrinde sich auch flächenmäßig ausdehnten, die Entstehung einer neuen Modalität statt. Wir bezeichnen diese neue Modalität als Orientierungsmodalität des magnocellularen Dunkel-On-Systems. Ihr Output informiert über das Vorhandensein von schwarzen Linienelementen. Im magnocellularen Hell-On-System entstand ebenso eine neue Modalität, die Orientierungsmodalität des magnocellularen Hell-Systems. Sie analysiert das Vorhandensein von weißen Linienelementen.
Da alle neuen Modalitäten sich im Verlauf der Evolution verselbständigten und sich auftrennten, gab es nach ihrer Aufspaltung so viele Retinaabbilder, wie es Orientierungssäulen gab. Jede Retinaabbildung war für einen Winkel und einen Nullpunktabstand zuständig. Gleichzeitig gab es eine Trennung in weiße und schwarze Linien. Es ist etwa so, als würde anstelle eines Fernsehgerätes 60 Fernsehgeräte nutzen, von denen die Hälfte die weißen Linienelemente anzeigt und die andere Hälfte die schwarzen. Hierbei würde jeder Fernseher nur für diejenigen Linienelemente des Ausgangsbildes zuständig sein, deren Orientierungswinkel in einem ganz bestimmten Winkelintervall mit 30 Grad Intervallbreite liegt. Der erste Fernseher würde also nur weiße Linienelemente anzeigen, deren Orientierungswinkel zwischen 0 Grad und 30 Grad liegt. Der Fernseher daneben würde weiße Linienelemente anzeigen, deren Winkel zwischen 30 und 60 Grad liegt. Jeder Nachbarfernseher würde das bevorzugte Winkelintervall um weitere 30 Grad erhöhen, so dass es 30 Fernseher für die weißen Linienelemente gäbe. Die übrigen 30 Fernseher würden für schwarze Linienelemente die gleiche Selektion aufweisen.
Hier muss daran erinnert werden, dass die mittlere Zellsäule, also die Orientierungssäule mit den Koordinaten x = 0 und y = 0 zuerst da war und die Modalität Weiß bzw. Schwarz detektierte. Ihr Output belegte ein eigenes Gebiet im sekundären Cortex.
Dadurch eroberten sich die Tiere, die dies hervorbrachten, eine neue Lebenswelt. Sie konnten nun in der Dämmerung Formen erkennen, auch wenn das schwache Mondlicht keine Farberkennung ermöglichte.
Später, als auch die Outputschichten der Farbmodule an Dicke gewannen, konnten diese Tiere auch am Tage Formen erkennen, die farbig waren.
So entwickelten sich das Helligkeitssehen und auch das Farbsehen zum Formensehen. Orientierungssäulen konnten im Sehfeld den Anstellwinkel einer Geraden erkennen, wenn diese einen beliebigen Grauton oder eine beliebige Farbe hatte. Es wurde sowohl der Anstellwinkel erkannt als auch der Grauton bzw. die Farbe, beides parallel und gleichzeitig.
Das Schönste daran war, dass man dies nicht erlernen musste. Sobald die Neuronen im Gehirn in den betreffenden Schichten voll ausgereift waren, konnte diese Leistung erbracht werden.
Lebewesen, bei denen die cortikale Reifung erst nach der Geburt stattfindet, haben dagegen noch die Reifungsphase zu durchlaufen, bevor sie Konturen von grauen oder farbigen Objekten erkennen können.
Monografie von Dr. rer. nat. Andreas Heinrich Malczan